









Практичне заняття № 6
ТЕМА: ФІЗІОЛОГІЯ СЕРЦЕВО – СУДИННОЇ СИСТЕМИ. ПОКАЗНИКИ. МЕХАНІЗМ. РЕГУЛЯЦІЯ.
Кровообіг — це рух крові у кровоносних судинах тіла за рахунок рушійної сили серця або пульсуючих судин. Кровообіг у людини був експериментально встановлений видатним англійським лікарем і вченим Уїльямом Гарвеєм (1628). Гарвей довів, що кров не виникає в печінці з їжі і не зникає в різних органах тіла у процесі їх життєдіяльності, як вважали раніше, а що вона відтікає від серця через артерії і повертається до нього венами, безперервно циркулюючи в колах кровообігу, багаторазово проходячи їх за годину. Це було винятковим відкриттям, і рік публікації книги Уїльяма Гарвея було визнано датою народження фізіології як експериментальної науки.
Системи циркуляції рідин організму забезпечують їх рух і виконання ними основних функцій: живлення, дихання, виділення, обміну інформацією між клітинами тощо.
У людини, як і в наземних тварин, наявне друге (мале) коло кровообігу. Основними чинниками, що зумовили його виникнення, стало дихання повітрям і сили гравітації. Серце нагнітає кров під тиском, достатнім для того, щоб вона пройшла через судини всього тіла і повернулась знову до серця. У міру переміщення крові судинами енергія її тиску витрачається на подолання опору цих судин, і тиск поступово знижується. Але в легенях (на відміну від зябр) мембрана розділяє рідину (кров) і газ (повітря). Повітря не здатне ззовні підпирати стінку легеневого капіляра і протидіяти високому тиску в ньому. Тому серце виштовхує кров у легені під низьким тиском (20-25 мм рт. ст.), достатнім лише для подолання опору легеневих судин. А пройшовши через мале коло, кров повинна знову надійти до серця і вийти з нього під високим тиском, необхідним для подолання опору судин великого кола кровообігу.
Що стосується гравітаційного чинника, то цей чинник, на відміну від водних істот, на наземні істоти і людину діє на повну силу, особливо на серцево-судинну систему. Для забезпечення нормального кровопостачання всіх органів і тканин тіла за цих умов потрібен вищий тиск крові в артеріальному руслі. Це можливо лише за умови розділення великого і малого кіл кровообігу.
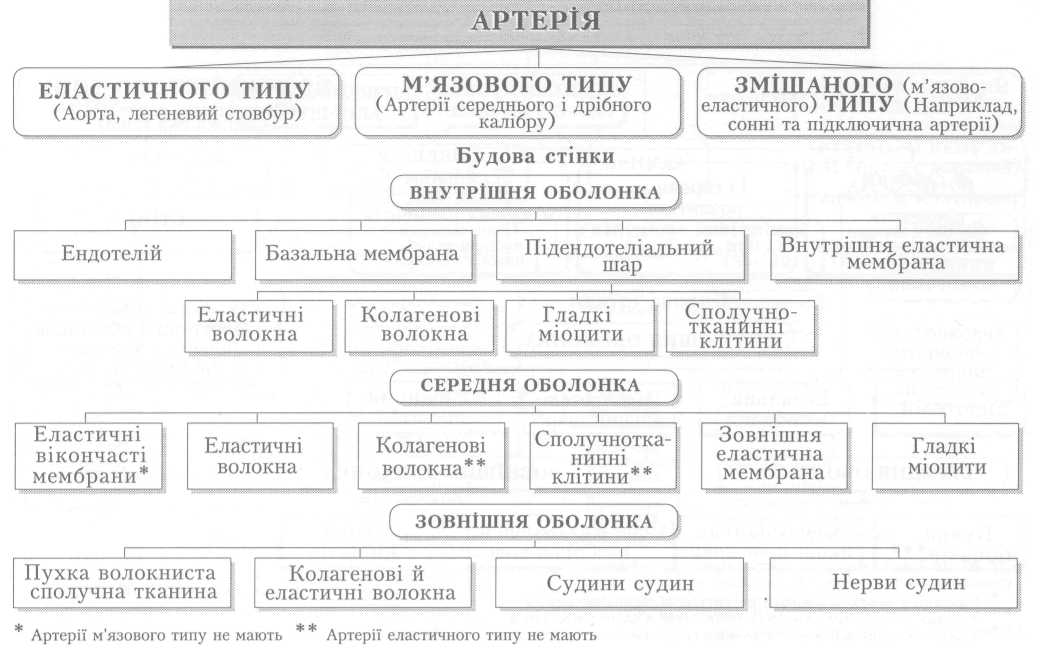
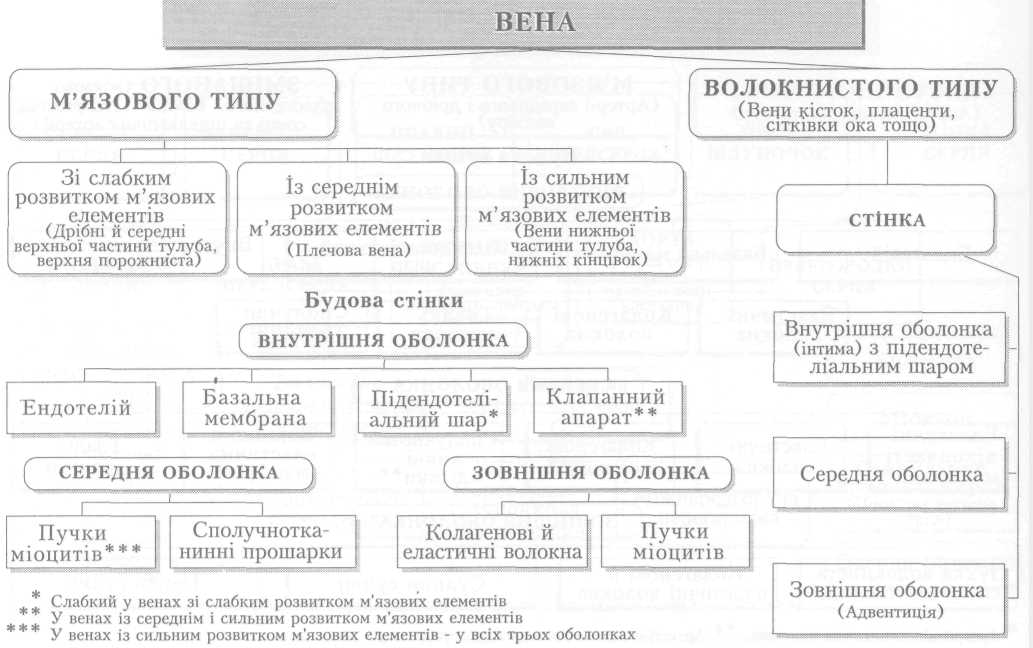
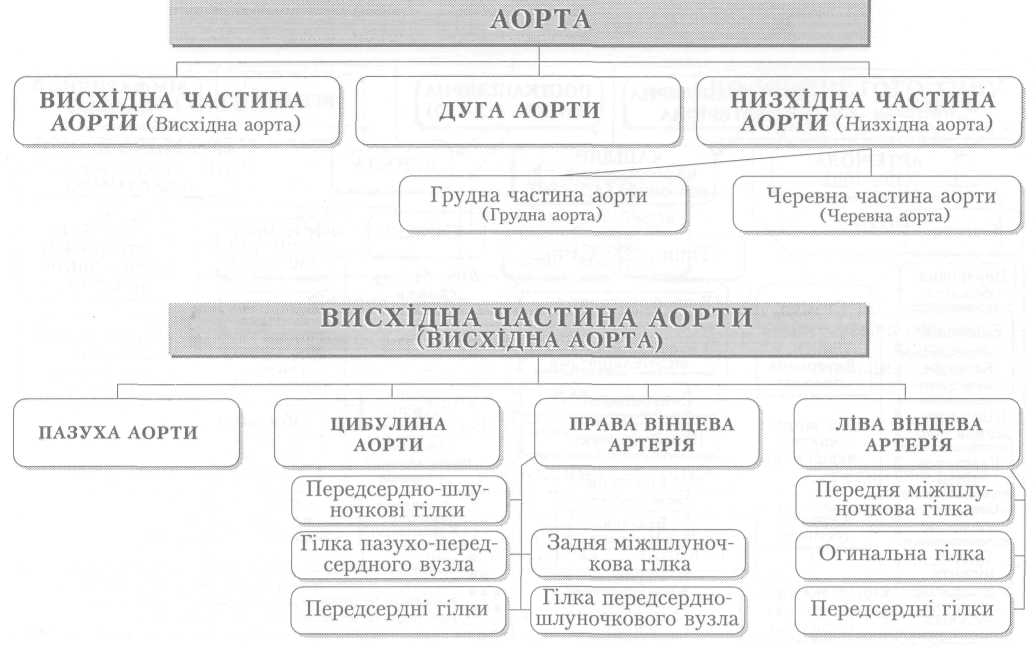
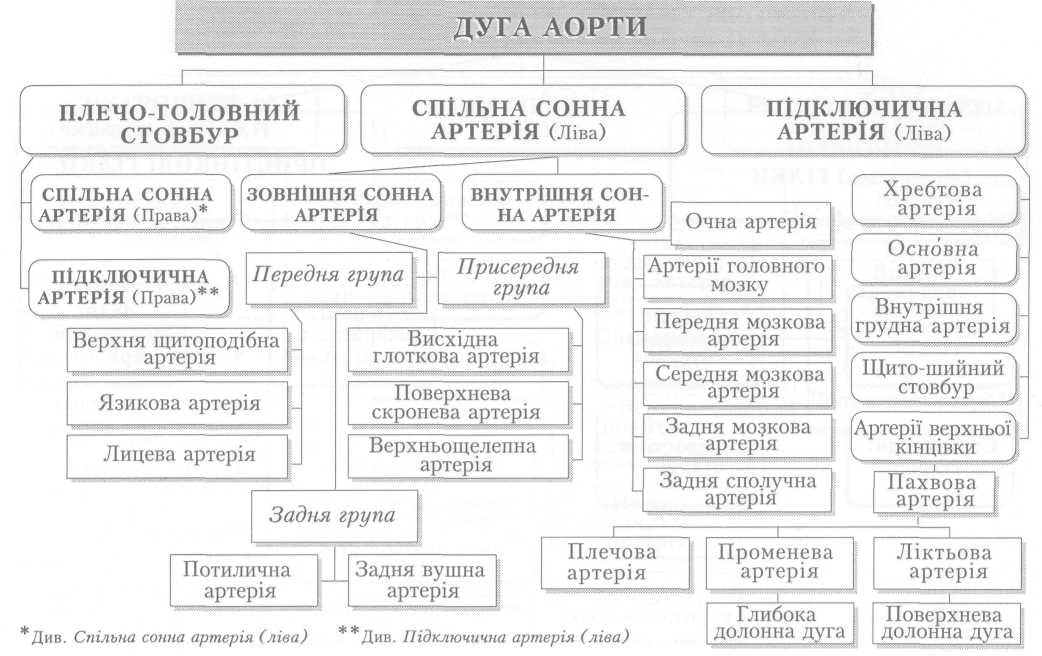
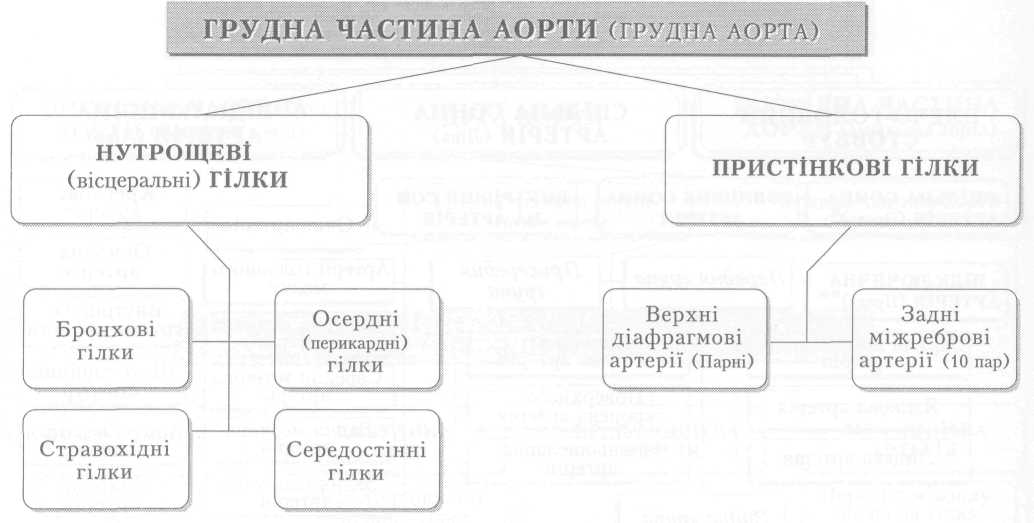
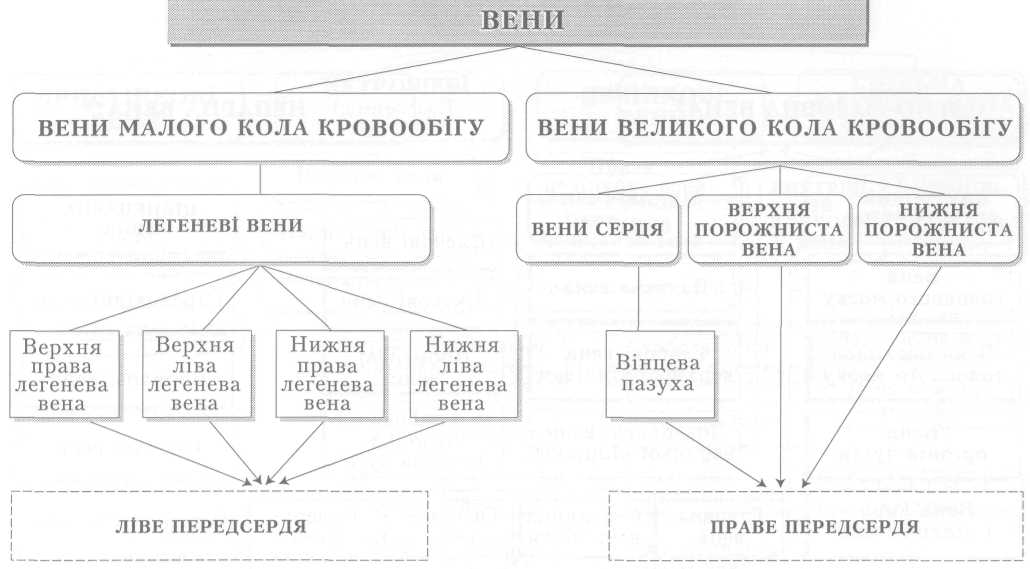
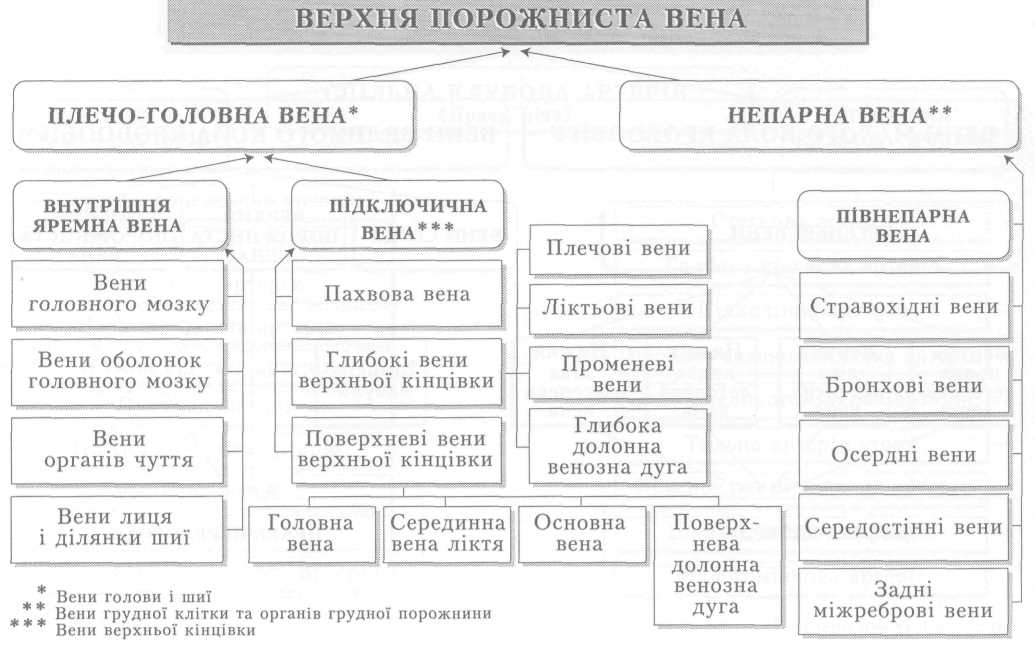
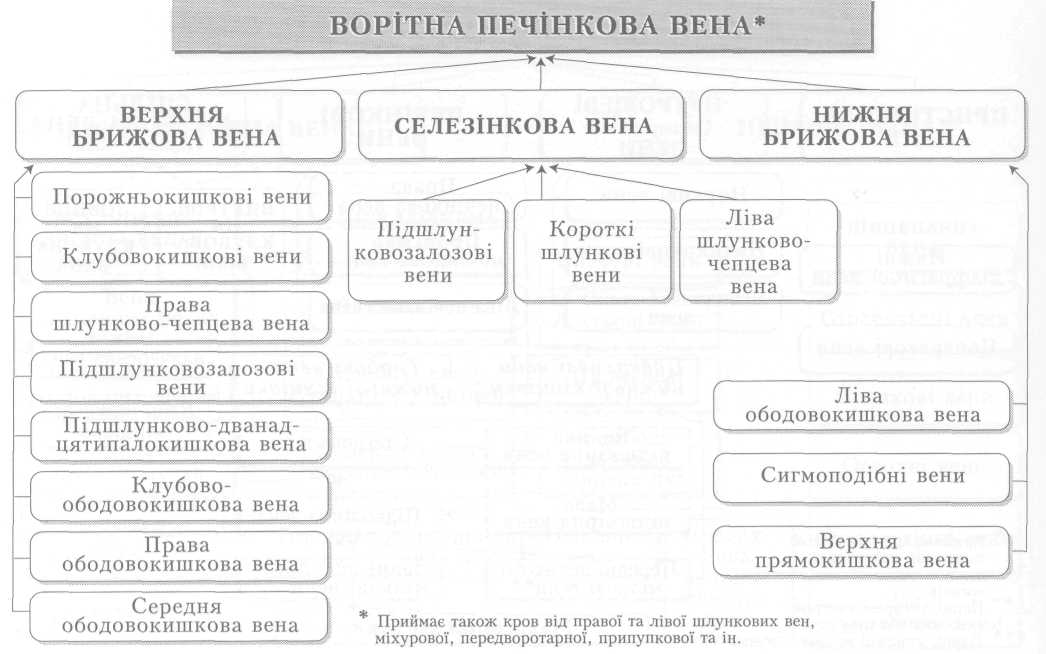
Велике коло кровообігу починається з лівого шлуночка серця, звідки виходить аорта. Через аорту та її гілки артеріальна кров розноситься по всьому тілу. Артерії різного калібру підходять до органів, розгалужуючись і закінчуючись капілярами, в яких кров віддає тканинам кисень, поживні речовини і забирає продукти обміну речовин. Далі капіляри сходяться у венули, дрібні, а згодом великі вени, які у людини зливаються в нижню (від тулуба й нижніх кінцівок) і верхню (від голови й верхніх кінцівок) порожнисті вени і несуть венозну кров до правого передсердя.
З правого шлуночка бере початок мале коло кровообігу. Від нього відходить легеневий стовбур і поділяється на праву та ліву легеневі артерії, якими венозна кров надходить до обох легень. Пройшовши через легеневі капіляри і обмінявшись газами з альвеолярним повітрям, уже артеріальна кров по чотирьох легеневих венах повертається до лівого передсердя, а звідти до лівого шлуночка і знову виштовхується в аорту.
С
Мал. 1. Будова серця (стрілками
позначено напрямок течії крові):
1 — дуга
аорти, 2 — легеневий
стовбур, 3 —
права і ліва легеневі артерії,
4 — верхня і нижня
порожнисті вени, 5 — легеневі
вени, 6 — праве
передсердя; 7 — правий
шлуночок, 8 — ліве
передсердя, 9 — лівий
шлуночок
БУДОВА СЕРЦЯ. ВЛАСТИВОСТІ СЕРЦЕВОГО М'ЯЗА
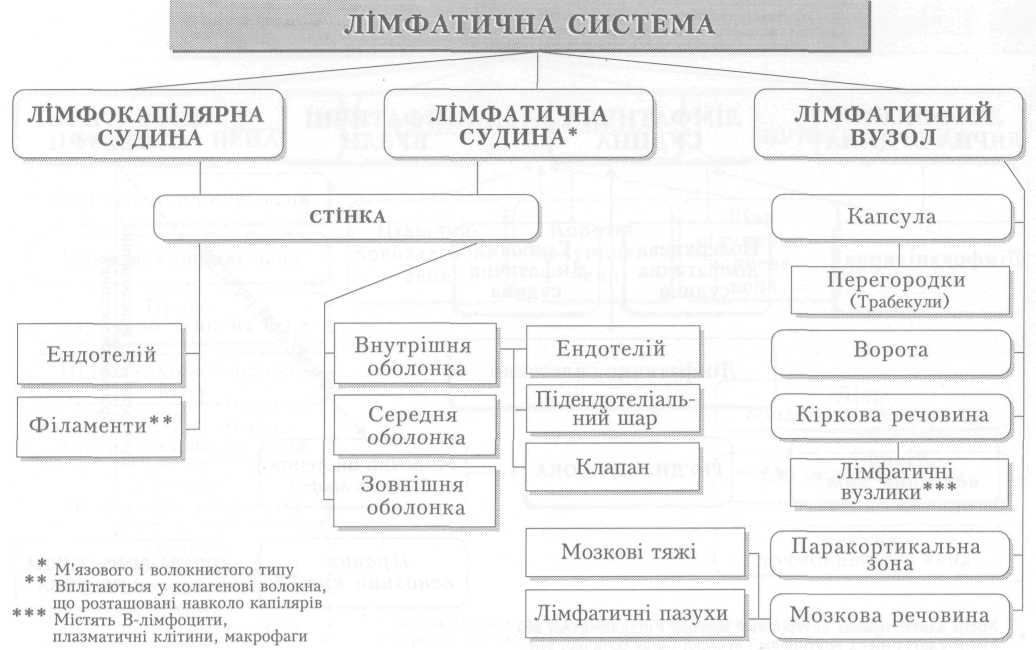
Серце — це порожнистий м'язовий орган, функція якого полягає у перекачуванні крові із судин з низьким тиском (вен) до системи судин високого тиску (артерій) і забезпеченні її плину в кровоносних судинах. Серце людини складається з чотирьох камер. Обидва передсердя так само, як і шлуночки, відокремлені одне від одного суцільною перегородкою, а передсердя і шлуночок кожної половини серця сполучаються між собою передсердно-шлуночковим (атріовентрикулярним) отвором з однойменним клапаном (мал. 1).
Стінка серця має неоднакову товщину, що залежить від навантаження на той чи інший відділ серця: найтоншою є стінка обох передсердь, найтовщою —лівого шлуночка (8-15 мм). Будова стінки в усіх відділах однакова. Це три шари: внутрішній — ендокард — вистеляє поверхню камер серця зсередини, утворює серцеві клапани і складається з шару ендотеліальних клітин та підстилаючої його підендокардіальної основи — сполучнотканинного шару з кровоносними судинами, нервами тощо. До нього прилягає найпотужніший шар стінки серця — міокард — м'язовий шар, що забезпечує основну функцію серця: перекачування крові з венозного до артеріального русла. Зовні його покриває сполучнотканинний шар — епікард. Крім того, існує ще четвертий шар — осердя (перикард), який охоплює все серце і утворює навколосерцеву сумку. Простір між осердям і епікардом заповнений невеликою кількістю серозної рідини. Остання під час скорочень серця зменшує його тертя з пристінковою поверхнею осердя.
Клапани серця виконують дуже важливу роль, забезпечуючи рух крові через серце тільки в одному напрямку. Клапани серця не мають м'язових елементів, відкриваються і закриваються пасивно за рахунок виникнення різниці тиску по обидва боки від них. Передсердно-шлуночкові клапани складаються зі стулок: лівий (мітральний, або двостулковий) — у лівій половині серця і правий (тристулковий) — у правій. Загальна поверхня стулок кожного клапана значно більша від площі отвору. Коли клапан закритий, стулки накладаються і частково перекривають одна одну. Кожна стулка — це еластична округло-трикутної форми складка-пелюстка ендокарда, яка основою фіксується до стінки отвору, а вершиною вільно звисає в бік шлуночка (мал. 2, а). Коли тиск крові в передсердях вищий, ніж у шлуночках, клапани відкриті, кров вільно надходить до шлуночків. Коли ж внаслідок скорочення шлуночків тиск у них зростає і стає вищим від передсердного, вільні краї пелюсток піднімаються догори, накладаються одна на одну і затуляють отвір. Вивертатися в бік передсердь стулкам не дають змоги сухожилкові струни, які одним кінцем прикріплені до вільного краю стулок, а другим до сосочкових м'язів, що виступають у порожнину шлуночка. Ці м'язи своїм скороченням підтримують постійне натягнення сухожильних ниток навіть під час скорочення серця, коли довжина шлуночка зменшується.
К
Мал. 2. Клапанний
апарат
серця:
а — лівий
передсердно-шлуночковий (мітральний,
двостулковий) клапан (вигляд збоку), б
— передсердно-шлуночкова
перегородка з клапанами серця (вигляд
згори)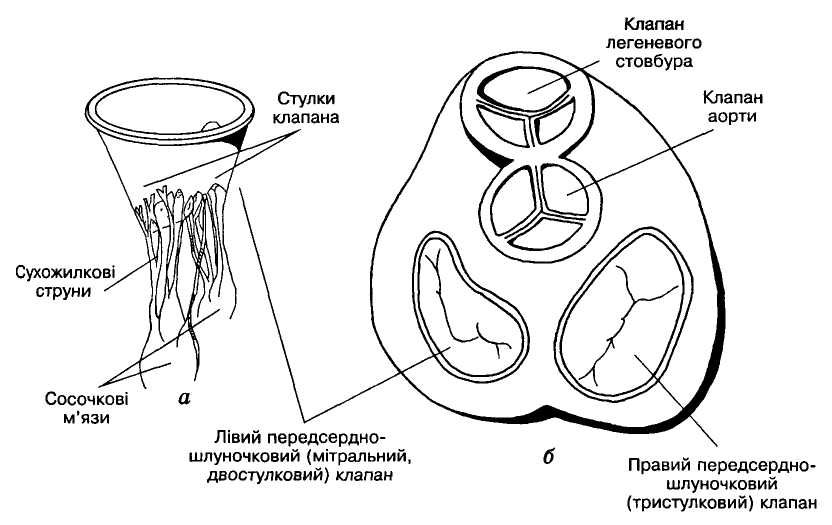
Клапани кожної половини серця відкриваються і закриваються в певній послідовності один за одним; періодично на короткий час вони бувають закриті одночасно обидва, і тоді шлуночки ізолюються від решти кровоносної системи. Проте ніколи обидва клапани не бувають відкриті одночасно. Вони забезпечують виключно односторонній рух крові через серце під час його скорочення. Щодо передсердь, то вони мають клапани лише на межі зі шлуночками. Між легеневими й порожнистими венами і відповідно лівим і правим передсердями клапанів немає, але є сфінктероподібні кільцеві м'язові утвори, які, скорочуючись одночасно з передсердями, перекривають вхід до них, запобігаючи виходу крові назад до вен під час скорочення передсердь.
Особливості серцевого м'яза. Серцевий м'яз — міокард — має унікальні особливості, за якими він істотно відрізняється як від посмугованих, так і від гладких м'язів. По-перше, це єдиний м'яз внутрішніх органів, що має поперечну посмугованість, а по-друге, це єдиний з посмугованих м'язів тіла, що є автономним, тобто незалежним від нашої волі. Виявлено ще деякі відмінності в його будові та властивостях.
За
будовою серцевий м'яз нагадує синцитій.
За
допомогою електронної мікроскопії
встановлено, що це не зовсім так. Клітини
міокарда —
серцеві
міоцити —
контактують
між собою переважно торцями за допомогою
вставних
дисків.
У
цих дисках мембрани обох клітин ідуть
паралельно, але не по прямій лінії, а
з
Мал. 3. Міжклітинні
сполучення у серцевому м'язі
А — пляма
злипання (сполучна пляма, щілинний
контакт, нексус), Б —
щілина вставного диска поза
сполучною плямою, В—Г
— вставний диск між двома
кардіоміоцитами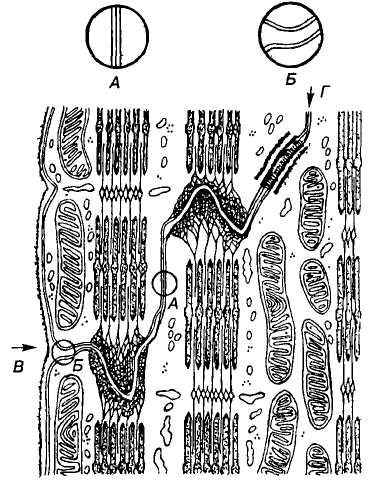
У дисках є плями злипання (сполучні плями, фасції злипання, щілинні контакти, нексуси), в яких ширина щілини між мембранами серцевих міоцитів не перевищує 4 нм (за межами нексуса 10-30 нм). У результаті проникність мембран у місці такого контакту для йонів у десятки разів вища, ніж за його межами. Завдяки цьому потенціал дії, а також збудження, що виникло в одній клітині, може охопити всю товщу міокарда, тобто серцевий м'яз є функціональним синцитієм, хоча морфологічно серцеві міоцити відокремлені один від одного мембранами.
Така особливість будови міокарда зумовила одну дуже важливу властивість, а саме: серце реагує на поодинокі подразнення за законом "усе або нічого". Тобто за сталих умов сила скорочення серцевого м'яза не залежить від сили подразнення, якщо вона досягла порогового чи понадпорогового рівня. Інакше кажучи, серце реагує скороченням лише на порогові й понадпорогові подразнення, причому скорочення завжди є максимальними. Серцевий м'яз завдяки наявності нексусів реагує як одна клітина. Якби міокард скорочувався не весь одночасно, то кров зі шлуночка не виштовхувалась би, а переливалась з однієї частини шлуночка до іншої.
Щ
Мал. 4. Рефрактерність
серця. Період рефрактерності: а—
абсолютної; б
— відносної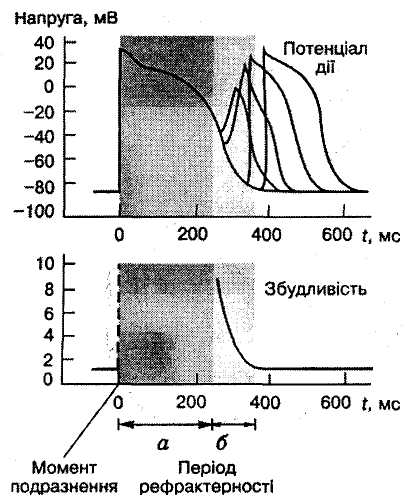
Та обставина, що серцевий м'яз протягом усього скорочення перебуває в стані рефрактерності, запобігає новому скороченню до закінчення попереднього, тобто тривалому тетанічному скороченню, що унеможливлювало б виконання серцем насосної функції. Крім того, така велика тривалість рефрактерного періоду запобігає рециркуляції збудження в міокарді, що також порушувало б нормальне чергування скорочення і розслаблення серця.
Відома висока невтомлюваність серцевого м'яза. Якщо людина не шкодить своєму серцю нездоровим способом життя, різноманітними перевантаженнями тощо, вона протягом усього життя не відчуває ніяких ознак втоми серця, яке за 60 років життя скорочується 2,3 млрд разів і перекачує понад 150 млн л крові. Така виняткова працездатність серця має кілька причин. Це, по-перше, ритмічність його роботи; по-друге, високий рівень кровопостачання міокарда і, нарешті, головна причина — висока ефективність метаболічних ферментів, здатних за 0,3 — 0,5 с між скороченнями серця повністю відновити біохімічний і енергетичний стан кардіоміоцитів. Значна тахікардія (понад 180 скорочень на 1 хв) є виснажливою навіть для нашого невтомного серця і, якщо вона не короткочасна, може призвести до патології.
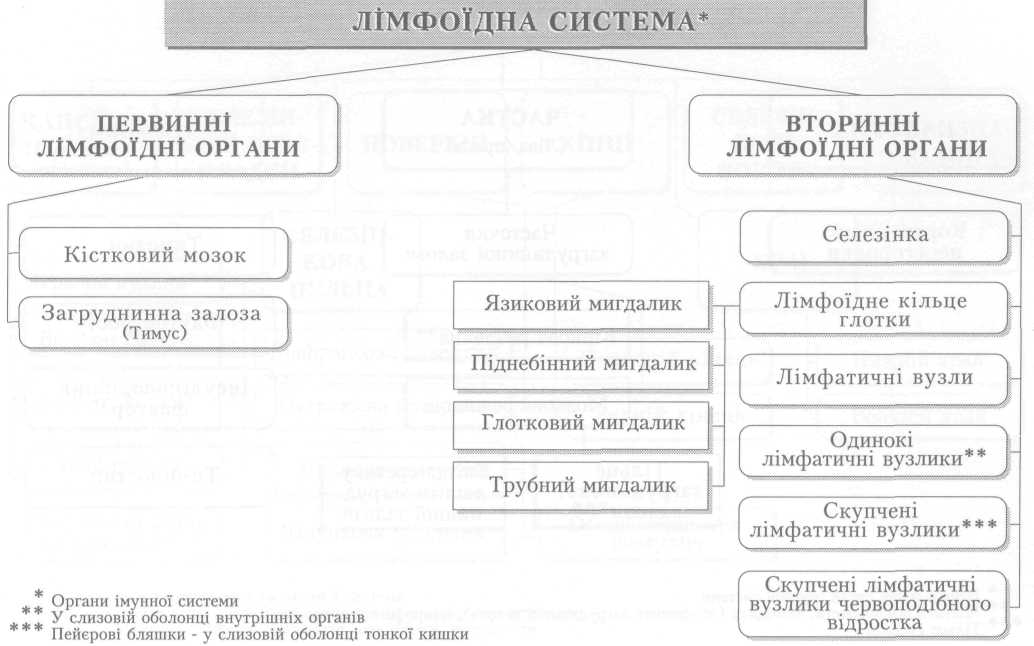
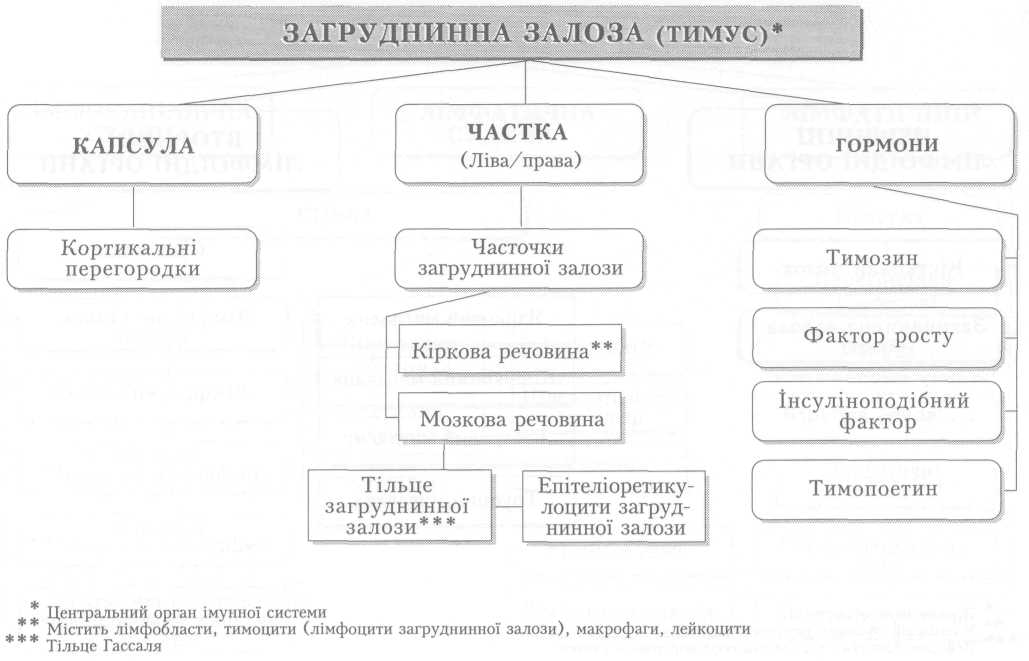
Автоматія (автоматизм) серця посідає особливе місце серед його властивостей. Це здатність ритмічно скорочуватись без будь-яких зовнішніх подразників, під впливом імпульсів, що виникають у самому серці. Автоматія властива усьому серцю, а також окремим його частинам, але не самому серцевому м'язу. Доказом автоматизму серця є факт ритмічних скорочень ізольованого серця різних тварин, а також людини.
Природа автоматії серця давно цікавила вчених. Досліди, відомі під назвою лігатур Станніуса, тобто ізолювання частин серця накладанням перев'язок, показали, що в серці є принаймні два центри автоматизму. Перший з них є провідним, що задає ритм скорочень усьому серцю, міститься у венозній пазусі (венозному синусі), а другий — на межі між передсердями і шлуночком, і він починає працювати лише після від'єднання першого центру. Серцевий м'яз без цих центрів не здатний до самостійних ритмічних скорочень.
Г
Мал. 5. Провідна
система серця: 1
— пазухо-передсердний вузол
(Кіса-Флека); 2 — передсердно-шлуночковий
вузол (Ашоффа-Тавари); 3 —
передсердно-шлуночкоиий
пучок (Гіса); 4 — ніжки
передсердно-шлуночкового пучка; 5
— серцеві провідні волокна
(волокна Пуркіньє); 6 — верхня
порожниста вена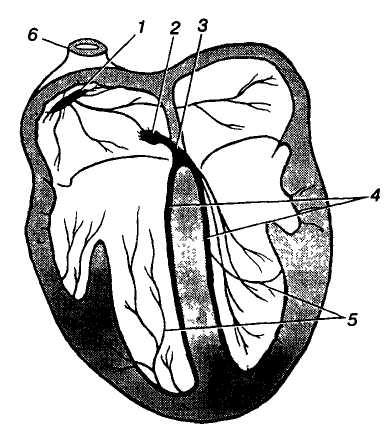
Ці факти переконливо свідчать, що автоматія серця має міогенне походження.
Автоматія серця зумовлена діяльністю провідної системи серця, яка у людини представлена скупченнями нетипових м'язових клітин — серцевих провідних міоцитів і м'язових волокон (мал. 5). Головним, або провідним, центром автоматії серця (водієм ритму, пейсмекером першого порядку) є пазухо-передсердний, або синусно-передсердний, вузол (вузол Кіса — Флека), розміщений під епікардом правого передсердя поблизу впадіння в нього верхньої порожнистої вени — у пазусі (синусі) порожнистих вен.
Від пазухо-передсердного вузла відходять кілька пучків до міокарда обох передсердь і до другого — передсердно-шлуночкового вузла, що є пейсмекером другого порядку, розміщеного під ендокардом між правим передсердям і шлуночком. Від нього відходить передсердно-шлуночковий пучок (пучок Гіса), який на початку міжшлуночкової перегородки поділяється на дві ніжки. Кожна з них йде уздовж міжшлуночкової перегородки під ендокардом у правому й лівому шлуночках до їхнього дна і далі, звертаючи на бокові стінки, віддає до клітин міокарда шлуночків тонкі волокна — серцеві провідні міоцити (волокна Пуркіньє). Передсердно-шлуночковий пучок та його розгалуження дістали назву пейсмекерів третього порядку.
Різні частини провідної системи мають неоднакову здатність до автоматизму; вона зменшується від пазухо-передсердного вузла до серцевих провідних волокон у шлуночках. Це явище має назву градієнта автоматії. За нормальних умов ритм серця задається пазухо-передсердним вузлом, якому підпорядковується провідна система. Проте у випадках, коли цей вузол не функціонує або збудження від нього не досягає передсердно-шлуночкового вузла, останній бере на себе функцію генерації ритму серця, частота скорочень при цьому значно зменшується. Передсердно-шлупочковий пучок, його ніжки і серцеві провідні волокна також можуть генерувати збудження, проте їх здатність до автоматизму є ще слабшою і виявляється лише у разі патології серця.
О
Мал. 6. Потенціали
дії (ПД) і повільна діастолічна
деполяризація (ПДД) клітин
пазухо-передсердного вузла (а) і міокарда
шлуночків (б)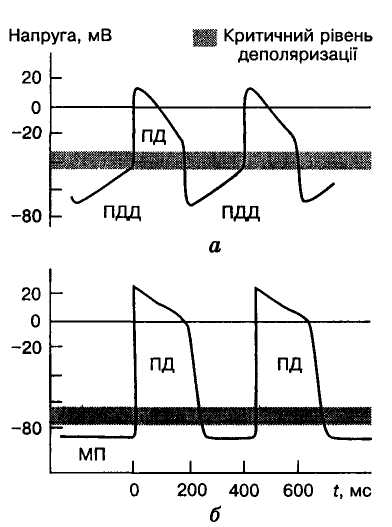
Тривала затримка проведення збудження — досить незвичне явище в фізіології збудливих систем, що довго не мало пояснення. Нині це явище пов'язують з морфофункціональними особливостями клітин передсердно-шлуночкового вузла: їхні численні тонкі відростки утворюють густу сітку, волокна якої зорієнтовані перпендикулярно до напрямку поширення збудження. Ця обставина й зумовлює різке зниження швидкості проведення і затримку збудження в передсердно-шлуночковому вузлі.
Що стосується механізму генерації ритмічних збуджень у серці, то їх пов'язують із функціональними особливостями клітин провідної системи серця. Усі клітини міокарда під час збудження генерують потенціали дії (ПД), а у клітин провідного центру автомата потенціалу дії передує нетривале і відносно невелике зменшення мембранного потенціалу (МП) — повільна діастолічна деполяризація (ПДД; мал. 6, а, б), виникнення якої зумовлене особливостями мембрани цих клітин та йонними струмами, що протікають крізь неї. Коли ПДД досягає порогового значення (близько 40 мВ), виникають потенціал дії і збудження, які через провідну систему поширюються на всі волокна міокарда, викликаючи скорочення його. Процес повторюється, зумовлюючи ритмічні скорочення серця.
Повільна діастолічна деполяризація властива не тільки клітинам пазухо-передсердного вузла. У більшості волокон провідної системи потенціалу дії серця передує ПДД, яка розвивається значно повільніше, ніж у клітинах пазухо-передсердного вузла, і тому не встигає досягти критичного рівня деполяризації. ПД в них виникає під впливом збудження в момент, коли воно надходить від пазухо-передсердного вузла, і неодночасність його виникнення в різних клітинах провідної системи зумовлена тривалістю проведення цього збудження до них.
РОБОТА СЕРЦЯ ТА ЇЇ ПРОЯВИ
Під час кожного скорочення серце виконує механічну роботу з виштовхування певного об'єму крові в аорту під певним тиском. Цю роботу (А) можна визначити кількісно як добуток сили на масу, в цьому разі тиску (Р) в аорті на об'єм крові, що виштовхується (Q):
А = Р • Q. (1)
Підрахунки показують, що в спокійному стані організму серце за одне скорочення виконує роботу близько 1 Дж, розвиваючи потужність 50 кВт/год.
Ритмічні скорочення серця супроводжуються певними механічними та електричними проявами, які несуть інформацію про роботу серця і мають певне діагностичне значення.
М
Мал. 7. Прояви
роботи серця. Запис електричної і
механічної активності серця: а
— електрокардіограма:
(PQRST — серцевий
цикл), б —
фонокардіограма: І—IV
— тони серця
Початок першого тону збігається із закриттям передсердно-шлуночкових клапанів на початку систоли шлуночків, а його продовження — з дрижанням сухожильних струн, що утримують стулки клапанів. Другий тон виникає в момент закриття клапанів аорти і легеневого стовбура і початку розслаблення шлуночків, коли тиск у них стає нижчим, ніж в аорті та легеневому стовбурі, і кров з цих судин рухається у зворотному напрямку, закриваючи клапани.
Таким чином, тривалість інтервалу між першим і другим тонами відповідає механічній систолі серця. На фонокардіограмі можна помітити також третій і четвертий тони. Третій тон зумовлений швидким припливом крові на початку наповнення шлуночків, а четвертий — скороченням передсердь. Ці два тони слабкі й на слух не сприймаються. Шуми серця між першим і другим тонами є ознакою вад серця: внаслідок деформації стулок герметичність клапанів порушується і кров, просочуючись крізь клапани у зворотному напрямку, спричинює ці шуми.
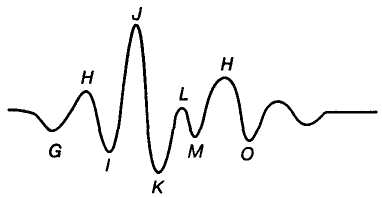
Б
Мал. 8. Балістокардіограма:
G, H, I,
J,
K — систолічні
хвилі, L, M,
H,
O — діастолічні
хвилі

Електричні прояви роботи серця. Електрокардіографія. Серце, як і будь-який м'яз, у процесі своєї діяльності продукує електричні струми. Кожному скороченню серця передує комплекс електричних коливань, який є сумарним потенціалом дії всіх волокон міокарда. Серце є генератором електричних потенціалів, а прилеглі до нього тканини передають ці потенціали на поверхню тіла. Проекція потенціалів на симетричні точки поверхні тіла є неоднаковою, і цю різницю потенціалів можна зареєструвати, приклавши до певних ділянок шкіри відвідні електроди і посиливши сигнали від них. Такий запис дістав назву електрокардіограми (ЕКГ) (див. мал. 7, а), а метод запису і подальшого аналізу потенціалів серця — електрокардіографії. Отже, ЕКГ — це запис різниці потенціалів між точками поверхні тіла, що відображає процеси збудження серцевого м'яза.
П
Мал. 9. Стандартні
відведення (/-///) електрокардіограми
(трикутник Ейнтховена)
На мал. 7, а наведено нормальну електрокардіограму людини в II стандартному відведенні (права рука — ліва нога). Зубці ЕКГ позначають латинськими літерами Р, Q, R, S і Т. Крім зубців на ЕКГ розрізняють також інтервали P—Q,Q—T,R — R. Зубець Р виникає в момент збудження передсердь. Ізоелектрична лінія P — Q відповідає переходу збудження на міокард шлуночків, її тривалість зумовлена затримкою збудження в передсердно-шлуночковому вузлі. Комплекс QRS відображає поширення збудження в міокарді шлуночків, інтервал S—Т означає, що обидва шлуночки цілком охоплені збудженням, зубець Т —процес виходу шлуночків зі етапу збудження. На мал. 10 поряд зі стандартною ЕКГ в тому самому масштабі часу наведено потенціал дії (ПД) кардіоміоцита, одержаний при внутрішньоклітинному відведенні за допомогою мікроелектрода. Виникає запитання, чому за дуже простої форми ПД кардіоміоцитів ЕКГ серця, що відводиться від поверхні тіла, має таку складну форму?
З
Мал. 10. Потенціал
дії кардіоміоцита (внутрішньоклітинне
відведення) (а) і
відповідний комплекс ЕКГ (б)
Мал. 11. Диференційна
гіпотеза походження електрокардіограми.
Різні форми ЕКГ (а, б)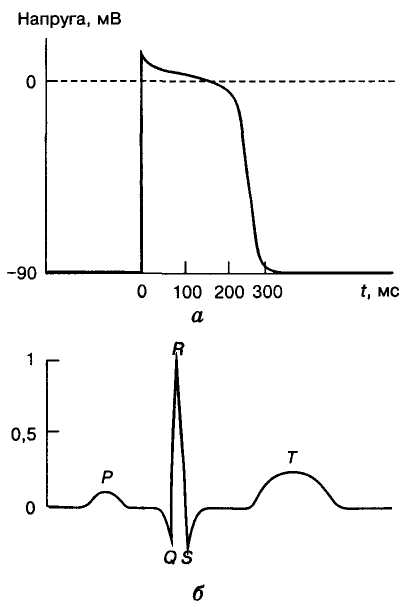

За іншою теорією — теорією рухомого диполя, збуджений кардіоміоцит є змінним диполем, параметри якого зображаються стрілкою певної довжини і напрямку — дипольним вектором, спрямованим від збудженої ділянки до незбудженої, тобто від мінуса до плюса. Алгебраїчна сума окремих векторів усіх кардіоміоцитів утворює інтегральний вектор серця. Оскільки в серці збудження розвивається за часом і поширюється в різних напрямках з неоднаковою швидкістю, то й дипольні та інтегральний вектори постійно змінюють свій напрямок і розмір у просторі.
Між параметрами інтегрального вектора і зубцями ЕКГ існує тісний зв'язок: розмір вектора прямо пропорційний амплітуді зубців ЕКГ і, коли на ЕКГ пишеться ізоелектрична лінія (сегменти P—Q, S — T i Т — Р), вектор стає нульовим.
Інформативність ЕКГ. Оскільки ЕКГ є записом електричних потенціалів, що генеруються серцем під час його збудження, то очевидно, що форма ЕКГ (амплітуда і форма зубців, тривалість інтервалів) надає інформацію про стан збудження серцевого м'яза, напрямок і швидкість поширення збудження і пов'язаний з цим процесом ритм скорочень серця. Так, за інтервалом R—R визначають частоту скорочень серця (ЧСС). В нормі у людини цей показник у стані спокою становить 60-80 скорочень за 1 хв. При ЧСС понад 90 за 1 хв говорять про тахікардію, а меншій за 60 — брадикардію. У добре тренованих спортсменів ЧСС у спокої може досягати 40 за 1 хв і менше.
А
Серед методів, які останнім часом використовують для дослідження функції серця в експерименті та клініці, слід назвати ехокардіографію і катетеризацію серця.