

Особенности формирования эксплицитной памяти
Уже отмечалось, что долговременная память делится на эксплицитную и имплицитную, в зависимости от участия сознания в процессе запоминания информации. Предполагается, что информация в эксплицитной памяти может храниться бесконечно долго, тогда как следы имплицитной памяти склонны угасать при отсутствии использования. Есть основание считать, что имплицитная память возникла в эволюционном развитии раньше, нежели эксплицитная.
Эксплицитная память (explicite — ясный, осознанный, англ.) связана с быстрым осознанным обучением. С ее помощью человек различает знакомые и незнакомые события. Многочисленные исследования подтвердили, что структуры, обеспечивающие функционирование этого типа памяти, находятся в височных областях мозга.
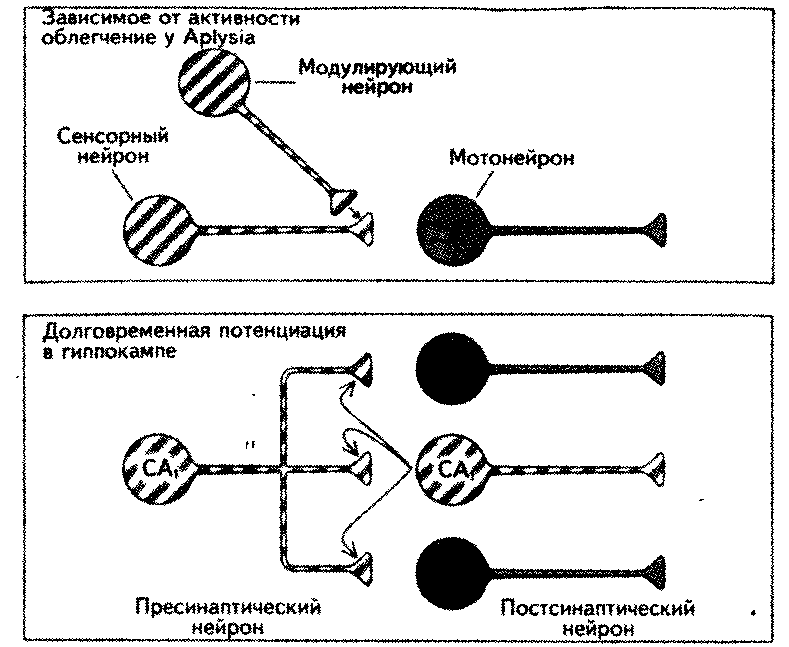
Д. Хебб (Hebb, 1949) сформулировал теоретически возможный механизм хранения информации. С его точки зрения, основой обучения может быть совпадение во времени активности пресинаптической и постсинаптической клеток. В дальнейшем такого типа связь между клетками стала называться синапсом Хебба. Сейчас считается, что эксплицитная память на нейрональ-ном уровне представлена пре-постсинаптическим совпадением активности нейрона (рис. 15.3). В 1986 г. Б. Вигстрем и X. Густафсон показали, что пре-постсинаптический ассоциативный механизм действует в гиппокампе (рис. 15.4) (Gustafsson, Wigstrom, 1990).
Рис. 15.3. Ассоциативные изменения силы связи между нервными клетками (Кандэл, Хо-ihc, 1992).
В этом случае при прохождении электрического импульса через терминаль
339
пресинаптического аксона происходит выход медиатора — глутамата — в си-наптическую щель. Глутамат активирует NMDA (М-метил-О-аспартат)-рецеп-торы на постсинаптической мембране, что приводит к запуску последовательности реакций, результатом которых является выработка оксида азота — недавно открытого медиатора нового типа. Молекулы этого вещества невелики и легко проходят сквозь мембрану постсинаптической клетки. Большее количество его инактивируется ферментами, а часть оксида азота, которая возвращается к пресинаптической терминали, возбуждает рецепторы кальциевых каналов в ней, способствуя дополнительному выбросу глутамата из везикул.
Вся последовательность событий повторяется снова и снова, вызывая долговременную потенциацию, если действие оксида азота сочетается с активностью пресинаптического нейрона. В основе долговременной потенци-ации в этом случае лежит сочетание двух независимых механизмов ассоциативного обучения: механизм Хебба с участием NMDA-рецепторов и зависимого от активности пресинаптического облегчения. Образование долговременных следов сопровождается увеличением числа пресинаптических терминалей (рис. 15.5).

Рис. 15.5. Долговременная потенциация сопровождается деполяризацией постсинаптической мембраны в результате активности HE-NMDA-рецепторных каналов. Деполяризация снимает блокаду NMDA-каналов магнием, что позволяет кальцию поступать в клетку через эти каналы. Кальций активирует зависимые от него киназы, и это приводит к индукции долговременной потенциации. Постсинаптическая клетка, по-видимому, выделяет ретроградный сигнал, способный проникать через мембрану пресинаптической клетки. Считается, что этот сигнал, которым, возможно, служит окись азота, действует на пресинаптическую тер-миналь, усиливая выделение нейромедиатора глутамата путем активации гуанилатциклазы или АДФ-рибозилтрансферазы (Кандэл, Хокинс, 1992).
340