

Тема 2. Основы общей генетики
Основные понятия генетики. Генетика – это наука о наследственности и изменчивости организмов.
► Наследственность – важнейшая особенность живых организмов, способность передавать свои свойства и функции от родителей к потомкам. Эта передача осуществляется с помощью генов.
Общие закономерности наследования систематизированы в рамках хромосомной теории наследственности - учении о локализации наследственных факторов в хромосомах клеток: преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом. Центральными являются понятия «хромосома», «генотип», «ген» и «аллель».
Хромосомная теория наследственности была сформулирована в 1902 г. Сэттоном и Бовери: хромосомы являются носителями генетической информации, определяющей наследственные свойства организма. У человека в каждой клетке имеется 46 хромосом, разделенных на 23 пары (рис. 2.1). Хромосомы, образующие пару, называются гомологичными хромосомами.
Д вадцать
две пары хромосом называются аутосомами,
их набор одинаков для особей мужского
и женского полов. Двадцать третья пара
- это пара, которая определяет пол и
соответственно различается по своей
структуре у мужчин и женщин: женщины
являются носителями двух Х-хромосом,
а мужчины - одной Х-
и одной
Y-хромосомы.
вадцать
две пары хромосом называются аутосомами,
их набор одинаков для особей мужского
и женского полов. Двадцать третья пара
- это пара, которая определяет пол и
соответственно различается по своей
структуре у мужчин и женщин: женщины
являются носителями двух Х-хромосом,
а мужчины - одной Х-
и одной
Y-хромосомы.
►Хромосомы (греч. chroma - цвет и soma - тело) - линейные структуры, в которые организованы гены в ядре клетки.
►Кариотип - хромосомный набор (число, размер, форма хромосом).
Половые клетки (гаметы) образуются при мейозе. В результате в каждой половой клетке остается только по одной гомологичной хромосоме из каждой пары, т. е. 23 хромосомы. Такой одинарный набор хромосом называется гаплоидным. При оплодотворении, когда сливаются мужская и женская половые клетки и образуется зигота, двойной набор – диплоидный восстанавливается. В зиготе и у организма, который из нее развивается, одна хромосома из каждой пары получена от отцовского организма, другая – от материнского.
► Ген – единица хранения, передачи и реализации наследственной информации. Представляет собой специфический участок (последовательность нуклеотидов) молекулы ДНК (дезоксирибонуклеиновая кислота), в структуре которого закодирована структура определенного полипептида (белка). Многие участки ДНК не кодируют белки, а выполняют регулирующие функции. В структуре генома человека только около 2% ДНК представляют последовательности, на основе которых идет синтез информационной РНК (рибонуклеиновая кислота) - процесс транскрипции, которая затем определяет последовательность аминокислот при синтезе белков - процесс трансляции. В настоящее время полагают, что в геноме человека имеется около 30-40 тыс. генов.
► Генотип - сумма всех генов организма, наследственная конституция организма, совокупность всех наследственных задатков данной клетки или организма, т.е. набор генов, организованных в хромосомный ряд. Генотип организма является результатом слияния двух гамет (яйцеклетки и оплодотворяющего ее спермия). В узком смысле генотип - совокупность аллелей гена или группа генов, контролирующих развитие и проявление анализируемого признака у данного организма.
►Фенотип - совокупность всех признаков и свойств организма, которые выявляются в процессе индивидуального развития в данных условиях. Понятие фенотип распространяется на любые признаки организма, начиная от первичных продуктов действия генов - молекул РНК и полипептидов и кончая особенностями внешнего строения, физиологических процессов, поведения и т. д. Фенотипическое проявление признаков всегда реализуется на основе взаимодействия генотипа с комплексом факторов внутренней и внешней среды.
►Аллели (греч. allenon - различные формы) - это альтернативные формы гена, определяющие альтернативные формы одного и того же признака.
В норме у каждого человека имеется два аллеля каждого гена - по одному аллелю на каждой из хромосом. В популяциях каждый ген может встречаться в виде множества аллелей, что обеспечивает определенный уровень генетического полиморфизма (например, три аллеля обусловливают существование четырех групп крови у человека) и комбинативной изменчивости (закон независимого наследования признаков).
З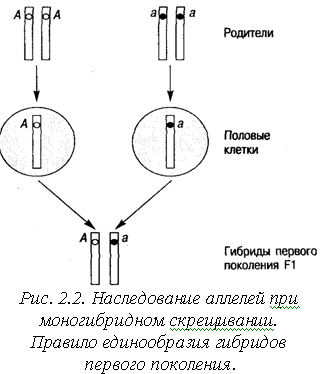 аконы
наследственности.
Основные законы наследственности были
описаны более века назад чешским монахом
Грегором Менделем (1822-1884), который
занимался селекционированием гороха,
и именно гороху, научной удаче и строгости
опытов Менделя мы обязаны открытием
основных законов наследуемости: закона
единообразия гибридов первого поколения,
закона расщепления и закона независимого
комбинирования.
аконы
наследственности.
Основные законы наследственности были
описаны более века назад чешским монахом
Грегором Менделем (1822-1884), который
занимался селекционированием гороха,
и именно гороху, научной удаче и строгости
опытов Менделя мы обязаны открытием
основных законов наследуемости: закона
единообразия гибридов первого поколения,
закона расщепления и закона независимого
комбинирования.
Аллели обозначают буквами латинского алфавита, при этом одни признаки Мендель назвал доминирующими (преобладающими) и обозначил их заглавными буквами - А, В, С и т. д., другие - рецессивными (уступающими, подавляемыми), которые обозначил строчными буквами - а, в, с и т. д. Поскольку каждая хромосома (носитель аллелей или генов) содержит лишь одну из двух аллелей, а гомологичные хромосомы всегда парные (одна отцовская, другая материнская), в диплоидных клетках всегда есть пара аллелей: АА, аа, Аа, ВВ, вв, Вв и т. д.
Особи и их клетки, имеющие в своих гомологичных хромосомах пару одинаковых аллелей (АА или аа), называются гомозиготными. Они могут образовывать только один тип половых клеток: либо гаметы с аллелью А, либо гаметы с аллелью а.
Особи, у которых в гомологичных хромосомах их клеток имеются и доминантный, и рецессивный гены Аа, называются гетерозиготными; при созревании половых клеток они образуют гаметы двух типов: гаметы с аллелем А и гаметы с аллелем а. У гетерозиготных организмов доминантная аллель А, проявляющаяся фенотипически, находится в одной хромосоме, а рецессивная аллель а, подавляемая доминантом, - в соответствующем участке (локусе) другой гомологичной хромосомы.
В случае гомозиготности каждая из пары аллелей отражает либо доминантное (АА), либо рецессивное (аа) состояние генов, которые в обоих случаях проявят свое действие.
В современной генетике приняты следующие условные обозначения при скрещивании: родительские формы - Р; полученные от скрещивания гибриды первого поколения - F1; гибриды второго поколения - F2, третьего - F3, и т. д. Само скрещивание двух особей обозначают знаком х (например: АА х аа).
Закон единообразия гибридов первого поколения. Скрещиваются организмы, различающиеся по одной паре признаков (моногибридное скрещивание). Пусть таким признаком будет цвет глаз.
На рис. 2.2 изображены пары аллелей на гомологичных хромосомах у родителей. У одного родителя это аллели А, соответственно его генотип для этих аллелей – АА. При данном генотипе цвет глаз карий. У другого родителя на обеих хромосомах находится аллель а (генотип аа), цвет глаз голубой.
В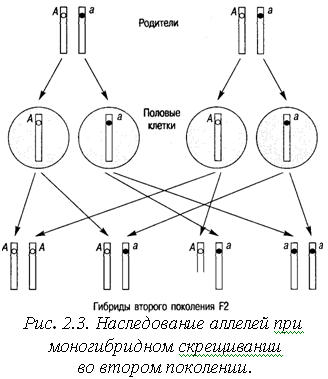 первом поколении (F1)
у потомства будет одинаковый генотип
Аа и
одинаковый фенотип – карие глаза. Аллель
а в
фенотипе не проявляется, присутствуя
в генотипе в «скрытом» виде. В данном
случае выполняется правило
(доминирования) единообразия гибридов
первого поколения: при
моногибридном скрещивании все потомство
в первом поколении характеризуется
единообразием по фенотипу и генотипу.
первом поколении (F1)
у потомства будет одинаковый генотип
Аа и
одинаковый фенотип – карие глаза. Аллель
а в
фенотипе не проявляется, присутствуя
в генотипе в «скрытом» виде. В данном
случае выполняется правило
(доминирования) единообразия гибридов
первого поколения: при
моногибридном скрещивании все потомство
в первом поколении характеризуется
единообразием по фенотипу и генотипу.
Закон расщепления. При скрещивании гибридов первого поколения (т.е. гетерозигот) между собой получается результат, изображенный на рис. 2.3. Половые клетки в процессе оплодотворения образуют гибриды второго поколения: гомозиготы АА, гетерозиготы Аа и гомозиготы аа.
Долю вероятности образования тех или иных генотипов удобно вычислять с помощью решетки Пенета (рис. 2.4). Вероятность образования гамет А или а одинакова для каждого родителя и составляет 0,5. Вероятность слияния двух гамет А равна произведению их вероятностей, т. е. 0,25, то же самое верно для генотипа аа. В результате во втором поколении наблюдается расщепление по генотипу в соотношении: 1АА : 2Аа:1аа.
Если принимать во внимание фенотип, то расщепление будет в соотношении 3:1, т. е. примерно у 3/4 потомков будут карие глаза (доминантный признак), и у 1/4 – голубые (рецессивный признак). Рецессивный признак проявляется только при условии, что рецессивный аллель оказывается в гомозиготном состоянии (аа).
Закон расщепления: у гибридов вт орого поколения в потомстве наблюдается расщепление по фенотипу доминантных и рецессивных признаков в соотношении 3:1.
Анализ семейных родословных показывает, что альбинизм у человека является рецессивной особенностью (рис. 2.5). Альбиносы гомозиготны по аллелю а (аа). У людей, гомозиготных по доминантному аллелю пигментации (АА) или гетерозиготных по этому гену (Аа), развивается нормальная пигментация. Брак альбиноса с нормальной особью ведет, как правило, к рождению нормальных детей, ибо в популяции большинство людей гомозитотны по аллелю А (АА). В случае брака между особями Аа и аа также рождается половина детей нормальных (Аа) и половина альбиносов (аа).
Я вление
полного доминирования наблюдается
далеко не всегда. Иногда наблюдается
промежуточный характер наследования
или неполное доминирование.
вление
полного доминирования наблюдается
далеко не всегда. Иногда наблюдается
промежуточный характер наследования
или неполное доминирование.
Закон независимого распределения. Скрещиваются особи, отличающиеся по двум парам аллелей и более. Каждая пара альтернативных признаков ведет себя в ряду поколений независимо друг от друга, в результате чего среди потомков первого поколения (у F2) в определенном соотношении появляются особи с новыми (по сравнению с родительскими) комбинациями признаков. Например, в случае полного доминирования при скрещивании исходных форм, различающихся по двум признакам, в следующем поколении (F2) выявляются особи с четырьмя фенотипами в соотношении 9:3:3:1. При этом два фенотипа имеют «родительские» сочетания признаков, а оставшиеся два - новые. Данный закон основан на независимом поведении (расщеплении) нескольких пар гомологичных хромосом. Так, при дигибридном скрещивании это приводит к образованию у гибридов первого поколения (F1) 4 типов гамет (АВ, Ав, аВ, ав), а после образования зигот - к закономерному расщеплению по генотипу и, соответственно, по фенотипу в следующем поколении (F2).
Парадоксально, но в современной науке огромное внимание уделяется не столько самому закону независимого распределения в его исходной формулировке, сколько исключениям из него. Закон независимого комбинирования не соблюдается в том случае, если гены, контролирующие изучаемые признаки, сцеплены, т.е. располагаются по соседству друг с другом на одной и той же хромосоме и передаются по наследству как связанная пара элементов, а не как отдельные элементы.
В случаях, когда наследуемость определенной пары генов не подчиняется этому закону Менделя, вероятнее всего эти гены наследуются вместе и, следовательно, располагаются на хромосе в непосредственной близости друг от друга. Зависимое наследование генов называется сцеплением, а статистический метод, используемый для анализа такого наследования, называется методом сцепления. Однако при определенных условиях закономерности наследования сцепленных генов нарушаются. Основная причина этих нарушений - явление кроссинговера, приводящего к перекомбинации генов. Биологическая основа рекомбинации заключается в том, что в процессе образования гамет гомологичные хромосомы, прежде чем разъединиться, обмениваются своими участками.
Законы Менделя в их классической форме действуют при наличии определенных условий:
1) гомозиготность исходных скрещиваемых форм;
2) образование гамет гибридов всех возможных типов в равных соотношениях (обеспечивается правильным течением мейоза; одинаковой жизнеспособностью гамет всех типов; равной вероятностью встречи любых гамет при оплодотворении);
3) одинаковая жизнеспособность зигот всех типов.
Знание и применение законов Менделя имеет огромное значение в медико-генетическом консультировании и определении генотипа фенотипически «здоровых» людей, родственники которых страдали наследственными заболеваниями, а также в выяснении степени риска развития этих заболеваний у родственников больных.

►Изменчивость – способность организмов приобретать новые признаки – различия в пределах вида. Выделяют две формы изменчивости: наследственную и модификационную (ненаследственную).
►Наследственная изменчивость – форма изменчивости, вызванная изменениями генотипа, которые могут быть связаны с мутационной либо комбинативной изменчивостью.
Гены время от времени подвергаются изменениям, которые получили название мутации.
►Мутации (лат. mutatio - изменение) - это внезапные, естественные или вызванные искусственно наследуемые изменения генетического материала, приводящие к изменению тех или иных фенотипических признаков организма. Имеют случайный характер и появляются спонтанно при воздействии определенных химических веществ, радиации, температуры и т. д. Предсказать появление той или иной мутации невозможно.
Возникшие мутации передаются потомкам, т. е. определяют наследственную изменчивость. Если мутация произошла в половой клетке, то у нее есть возможность передаться потомкам, т. е. быть унаследованной. Если мутация произошла в соматической клетке, то она передается только тем клеткам, которые возникают из этой соматической клетки. Такие мутации называются соматическими, они не передаются по наследству.
►Наследственная изменчивость (мутационная или генотипическая) связана с изменением генотипа особи, поэтому возникающие изменения наследуются. Она является материалом для естественного отбора. Дарвин назвал эту наследственность неопределенной. Основой наследственной изменчивости являются мутации - внезапные скачкообразные и ненаправленные изменения исходной формы. Они ведут к появлению у живых организмов качественно новых наследственных признаков и свойств, которых ранее в природе не существовало. Источник наследственной изменчивости - мутационный процесс.
Различают несколько типов мутаций: геномные, хромосомные и генные (см. главу 5).
Геномные мутации (полиплоидия и анеуплоидия) - это изменения числа хромосом. Полиплоидия - это кратное увеличение гаплоидного набора хромосом (Зn, 4n, и т.д.). Чаще всего полиплоидия образуется при нарушении расхождения хромосом к полюсам клетки в мейозе или митозе под действием мутагенных факторов. Она широко распространена у растений и крайне редко встречается у животных.
Анеуплоидия - увеличение или уменьшение числа хромосом по отдельным парам. Она возникает при нерасхождении хромосом в мейозе или хроматид в митозе. Анеуплоиды встречаются у растений и животных и характеризуются низкой жизнеспособностью.
Хромосомные мутации - это изменения структуры хромосом. Различают следующие виды хромосомных мутаций:
Дефишенсия - потеря концевых участков хромосом.
Делеции - выпадение участка плеча хромосом.
Дупликация - повторение набора генов в определенном участке хромосомы.
Инверсия - поворот участка хромосом на 180°.
Транслокация - перенос участка к другому концу той же хромосомы либо к другой, негомологичной хромосоме.
Генные мутации - изменения нуклеотидной последовательности молекулы ДНК (гена). Их результат - изменение последовательности аминокислот в полипелтидной цепи, и появление белка с новыми свойствами. Большая часть генных мутаций фенотипически не проявляется, поскольку они рецессивны.
Цитоплазматические мутации связаны с изменениями органоидов цитоплазмы, содержащих ДНК (митохондрии и пластиды). Эти мутации наследуются по материнской линии, т.к. зигота при оплодотворении всю цитоплазму получает от яйцеклетки. Пример: пестролистность растений связана с мутациями в хлоропластах.
►Комбинативная изменчивость определяется характером полового процесса и проявляется уже на стадии образования половых клеток - возникают новые генотипы из-за новых комбинаций генов. В каждой половой клетке (гамете) представлена только одна гомологичная хромосома из каждой пары. Хромосомы попадают в гамету абсолютно случайным образом, поэтому половые клетки одного человека могут довольно сильно отличаться по набору генов в хромосомах. Еще более важная стадия для возникновения комбинативной изменчивости – это оплодотворение, после которого у вновь возникшего организма 50% генов унаследовано от одного родителя, а 50% – от другого.
►Модификационная изменчивость – форма изменчивости, не связанная с изменениями генотипа и вызванная влиянием среды на развивающийся организм.
Наличие модификационной изменчивости очень важно для понимания сущности наследования. Наследуются не признаки. Можно взять организмы с абсолютно одинаковым генотипом, например, вырастить черенки от одного и того же растения, но поместить их при этом в разные условия (освещенность, влажность, минеральное питание) и получить достаточно сильно отличающиеся растения с разными признаками (рост, урожайность, форма листьев и т. п.).
Для описания реально сформировавшихся признаков организма используют понятие фенотип.
Фенотипы не наследуются, а формируются в течение жизни; они - продукт чрезвычайно сложного взаимодействия генотипа и среды.
►Фенотип – это весь комплекс реально возникших признаков организма. Фенотип формируется как результат взаимодействия генотипа и влияний среды в ходе развития организма. Фенотипы не наследуются, а формируются в течение жизни; они - продукт чрезвычайно сложного взаимодействия генотипа и среды. Таким образом, сущность наследования заключается не в наследовании признака, а в способности генотипа в результате взаимодействия с условиями развития давать определенный фенотип.
Важно отметить, что существуют единичные признаки, фенотип которых полностью определяется их генетическими механизмами. Примеры таких признаков - полидактилия (наличие добавочного пальца) или группа крови человека. Однако подобных признаков совсем немного, и за очень редким исключением фенотип признака определяется совместным влиянием генотипа и среды, в которой этот генотип существует.
Модификации не передаются по наследству. Обычно это положение почему-то с трудом принимается. Кажется, что если, скажем, родители на протяжении нескольких поколений тренируются в поднятии тяжестей и обладают развитой мускулатурой, то эти свойства должны обязательно передаться детям. Между тем это типичная модификация, а тренировки – это и есть то воздействие среды, которое повлияло на развитие признака. Никаких изменений генотипа при модификации не происходит, и приобретенные в результате модификации признаки не наследуются.
Для характеристики пределов модификационной изменчивости существуют понятия норма (диапазон) реакции. Некоторые признаки у человека невозможно изменить за счет средовых влияний, например группу крови, пол, цвет глаз. Другие, напротив, очень чувствительны к воздействию среды. К примеру, в результате длительного пребывания на солнце цвет кожи становится значительно темнее, а волосы, наоборот, светлеют. На вес человека сильно влияют характер питания, болезни, наличие вредных привычек, стресс, образ жизни.
Средовые воздействия могут приводить не только к количественным, но и к качественным изменениям фенотипа. У некоторых видов примулы при содержании при пониженной температуре (15-20°С) появляются цветы красного цвета, если же растения поместить во влажную среду при температуре 30°С, то образуются белые цветки.
Причем, хотя норма реакции характеризует ненаследственную форму изменчивости (модификационную изменчивость), она тоже определяется генотипом. Это положение достаточно важно – норма реакции зависит от генотипа. Одно и то же воздействие среды у одного генотипа может привести к сильному изменению признака и никак не повлиять на другой.
Общее в понятиях нормы и диапазона реакции заключается в следующем. Нормой (диапазоном) реакции данного генотипа называется система, описывающая множество фенотипов, существование которых потенциально возможно в том случае, если данный генотип будет находиться во взаимодействии с определенными средами. Понятия и нормы, и диапазона реакции предполагают, что каждый генотип ассоциируется с определенным, характерным для него, рядом фенотипов, формирующихся в разных средах. Упрощенно понятие нормы (диапазона) реакции можно представить следующим образом:

Каждому генотипу соответствует своя определенная норма (диапазон) реакции.

Различия в понятиях нормы и диапазона реакции состоят в следующем. Рассмотрим гипотетический пример, касающийся фенотипического признака, который отражает какие-то специфические способности. Предположим, существует 4 генотипа (1, 2, 3, 4), и все эти генотипы могут быть одновременно помещены в разные типы сред, отличающиеся друг от друга по уровню разнообразия и обогащенности. Схематически эта ситуация отражена на рис. 2.6.а.
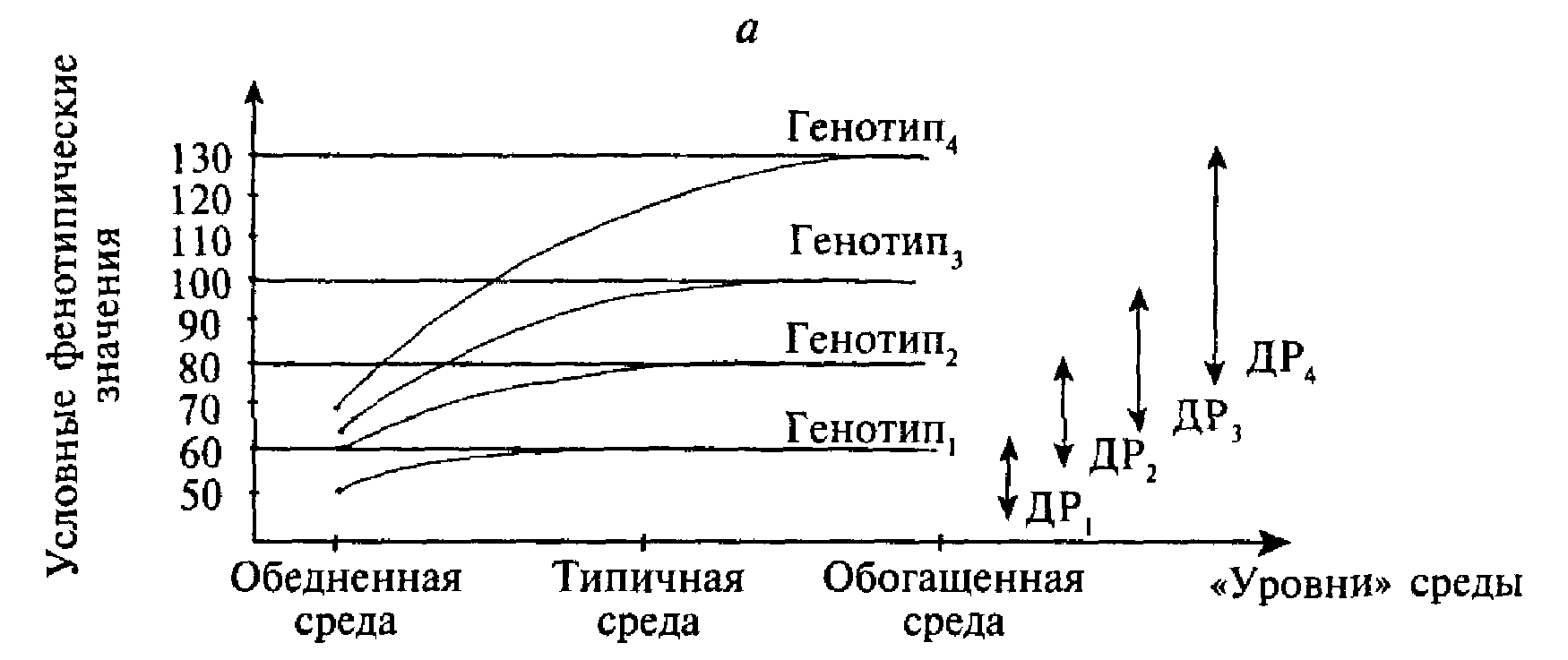

Рис. 2.6. Схематическое изображение зависимости фенотипических значений от типов генотипа и среды, в которой данный генотип развивается.
а - схематическая иллюстрация понятия «диапазон реакции» (ДР). Диапазон реакции обозначает разницу между фенотипическими значениями определенного генотипа в разных типах сред.
б - схематическая иллюстрация понятия «норма реакции». Один и тот же генотип (Генотип5 и Генотип6) в разной среде дает разные фенотипы.
По оси абсцисс отложены «уровни» среды, а по оси ординат — условные фенотипические значения. Согласно рисунку, в обедненной среде разброс фенотипических значений относительно мал, и четыре генотипа проявляются в фенотипах, мало отличающихся друг от друга. Разброс фенотипических значений существенно возрастает в типичной среде и достигает максимума в среде обогащенной. Разница между значениями данного генотипа в обедненной и обогащенной средах называется диапазоном реакции этого генотипа.
Понятие диапазона реакции подразумевает сохранение рангов фенотипических значений генотипов в разных средовых условиях. Например, Генотип1, ассоциируется с низкими фенотипическими значениями и в обедненной, и в обогащенной средах, в то время как Генотип4 является наиболее «процветающим» в любой среде. Соответственно, диапазон реакции Генотипа1 - наименьший, а диапазон реакции Генотипа4 - наибольший. Иными словами, основным допущением при интерпретации понятия «диапазон реакции» служит следующее предположение: существующие генотипы отличаются друг от друга таким образом, что фенотипические преимущества каждого из этих генотипов постоянны, а фенотипические различия, ассоциируемые с каждым из генотипов, становятся все более заметны по мере того, как среда становится все более благоприятной для развития данного фенотипического признака. Если взять в качестве примера математические способности, то носители Генотипа4 будут демонстрировать наивысшие значения как в обедненной, так и в обогащенной среде, причем чем благоприятнее среда, тем выше уровень математических достижений. Напротив, носители Генотипа1, будут иметь наименьшие фенотипические значения в любой среде, а фенотипические изменения, характеризующие этот фенотип при переходе из одних средовых условий в другие, будут незначительны.
Добавим к изучаемым нами генотипам два новых - Генотип5 и Генотип6 (рис. 2.6.б). Оказывается, что поведение этих двух генотипов в разных средах не соответствует ожиданиям о сохранении ранговых мест фенотипических выражений разных генотипов в варьирующих средовых условиях. Как показано на рис. 2.6.б, максимальное фенотипическое значение Генотипа5 наблюдается в типичной среде, в то время как обогащенная среда не является благоприятной для этого генотипа - его фенотипическое значение уменьшается. В качестве возможной иллюстрации данного феномена может быть использован хорошо известный из психологии развития факт: излишняя когнитивная стимуляция многих (но не всех) младенцев часто приводит не к оптимизации, а к расстройству их познавательной деятельности.
Генотип6, напротив, на переход от обедненной к типичной среде никак не реагирует, его фенотипическое значение остается неизменным. Однако ситуация существенно меняется при изменении средовых условий на обогащенные: фенотипическое значение Генотипа6 резко и линейно возрастает. Примером подобной ситуации может служить развитие музыкальных способностей, поскольку ребенок, основываясь на своих природных задатках, должен овладеть мастерством, для обучения которому ему необходимо находиться в обогащенной среде, в то время, как и обедненная, и типичная среды таких условий не дают.
Таким образом, несколько упрощая ситуацию, можно сказать, что понятие нормы реакции - более общее понятие, поскольку, используя его, исследователь не должен делать никаких предположений о сохранении рангов фенотипов в разных средах. Для понятия же диапазона реакции допущение об определенном ранговом порядке фенотипов (и, соответственно, генотипов) в контексте разных средовых условий является критическим. В силу большей широты понятия нормы реакции далее в учебнике будет использоваться именно это понятие.
Типы наследования и экспрессия генов. Носители генетической информации - гены - это участки хромосомной ДНК, в которых закодирована аминокислотная последовательность полипептидов или белков. Белок нормально функционирует лишь в том случае, если его молекула надлежащим образом свернута, а для этого правильные аминокислоты должны располагаться в ней в правильной последовательности.
Как было отмечено выше, генной мутацией называют изменение одного из многочисленных нуклеотидов ДНК - выпадение какого-нибудь нуклеотида, добавление нового нуклеотида или превращение одного нуклеотида в другой, что в свою очередь приводит к изменению белка, синтезируемого по инструкциям, содержащимся в данном гене. Мутация может сделать синтез соответствующего белка вообще невозможным или может привести к тому, что клетка начнет вырабатывать белок с иной аминокислотной последовательностью. Такой измененный белок будет выполнять свою функцию в организме менее эффективно. Серьезность последствий, к которым приведет мутация, зависит от того, как сильно она изменит белок, а также от того, насколько важен данный белок для организма.
В XX в. стало ясно, что некоторые врожденные нарушения есть не что иное, как фенотипическая экспрессия мутантных генов. Рассмотрим некоторые типы наследования, которые остались неизвестными Менделю; речь пойдет главным образом о примерах, касающихся человека. Мы узнаем также, как эти типы наследования связаны с тем фактом, что гены экспрессируются (проявляются) через свои продукты, т.е. через белки, кодируемые генной ДНК.
Летальные гены. Почему один ген является доминантным, а другой рецессивным? Мутантная ДНК, не кодирующая никакой белок или кодирующая белок, лишенный активности, не будет экспрессироваться, т. е. представляет собой рецессивный ген. У гетерозиготных индивидуумов нормальный ген направляет синтез нормального белка, а рецессивный ген никакого вклада в синтез функционального белка не вносит. Такой индивидуум имеет нормальный фенотип; нормальный ген в этом случае доминирует. У индивидуума, гомозиготного по данному рецессивному гену, соответствующий белок не образуется, а поэтому нормальная экспрессия данного признака невозможна. Рецессивный фенотип - это отсутствие нормального признака. Так, карликовость - результат «отсутствия ростовых веществ».
Если белок, о котором идет речь, необходим для жизни данного организма, то организм, неспособный образовать активную форму этого белка, обречен на гибель, и дефектный ген называется в этом случае летальным геном. Доминантные летальные гены существуют, но в большинстве случаев они быстро элиминируются, потому что вызывают гибель несущего их организма. Рецессивные же летальные гены могут и не причинять вреда гетерозиготному индивидууму; поэтому они могут передаваться следующим поколениям и распространяться в популяции весьма широко. Подсчитано, что в среднем человек гетерозиготен приблизительно по 30 летальным рецессивным генам. Это больше, чем у многих других организмов, и отчасти именно этим объясняется тот факт, что у людей среди потомков от браков между близкими родственниками процент летальных наследственных признаков выше, нежели при близкородственных скрещиваниях у большинства других видов.
Если одна копия «нормального» гена не обеспечивает образования его белкового продукта в количестве, достаточном для нормального функционирования организма, то по данному гену наблюдается неполное доминирование. В таких случаях гетерозигота отличается по фенотипу от обеих гомозигот. У человека, например, имеется летальный ген, обусловливающий в гетерозиготном состоянии брахидактилию, т.е. укорочение средней фаланги пальцев; палец при этом выглядит так, как будто в нем не три, а две фаланги. У гомозигот этот ген является причиной аномального развития скелета. Дети, гомозиготные по этому гену, рождаются без пальцев и с другими нарушениями в развитии скелета, приводящими к смерти в раннем возрасте.
В тех случаях, когда брахидактилией страдают оба родителя, можно ожидать, что из каждых четырех детей один будет гомозиготен по этому летальному гену и умрет в раннем детстве, два будут гетерозиготны, т.е. будут страдать брахидактилией, и один будет нормальным.
Широко известный ген человека, который в гомозиготном состоянии часто оказывается летальным, ответствен за серповидноклеточную анемию. Соответствующий нормальный ген кодирует одну из полипептидных цепей (бета-цепь) гемоглобина - белка, функция которого заключается в переносе кислорода; этот белок находится в эритроцитах и обусловливает их красный цвет. У мутантного гена один из нуклеотидов, присутствующих в нормальной ДНК, заменен другим, в результате чего и в бета-цепи гемоглобина одна из аминокислот заменена другой.
Это изменение, казалось бы столь ничтожное, влечет за собой роковые последствия. При аномально низком содержании кислорода молекулы гемоглобина в эритроцитах, содержащих гемоглобин S, агрегируют, образуя волокна. Вследствие этого эритроциты деформируются, приобретая серповидную форму. Такие эритроциты застревают в мелких кровеносных сосудах, и кровоснабжение участков, обслуживаемых этими сосудами, нарушается. Нарушение кровоснабжения вызывает разнообразные симптомы: чувство усталости, судороги, раздражительность.
Ген серповидноклеточности и соответствующий нормальный ген кодоминантны, так что у гетерозиготных индивидуумов синтезируются и нормальные и мутантные бета-цепи. Эритроциты таких людей приобретают серповидную форму лишь при крайне низком содержании кислорода, например, на больших высотах. Без специальных тестов такие гетерозиготные индивидуумы могут и не знать о том, что они являются носителями гена серповидноклеточности. Гомозиготные индивидуумы страдают гораздо сильнее, потому что у них дефектны все бета-цепи.
Индивидуумов, гетерозиготных по тому или иному генетическому нарушению, называют носителями, а гомозиготных - больными. Про людей, гетерозиготных по гену серповидноклеточности, иногда говорят, что они больны серповидноклеточной анемией. Это неверно, ибо это предполагает, что носитель данного гена менее жизнеспособен, нежели нормальный гомозиготный индивидуум. Между тем в действительности это обычно не так.
Около 90% индивидуумов, гомозиготных по гену серповидноклеточности, умирает в раннем возрасте. Можно было бы ожидать, что такой летальный ген будет быстро элиминироваться естественным отбором, поскольку гомозиготы по этому гену умирают слишком рано и не могут передать его потомству. Однако в тропической Африке на весьма обширных пространствах от 20 до 40% людей гетерозиготны по этому гену. В пятидесятых годах исследователи обратили внимание на один примечательный факт: оказалось, что эти люди живут в местах, где распространена особо вирулентная форма малярии.
Наличие хотя бы одной копии серповидноклеточности делает человека более устойчивым к малярии. Эритроциты, содержащие гемоглобин S, приобретают серповидную форму легче, если они заражены малярийным плазмодием. Когда эритроцит деформируется, находящийся в нем паразит погибает. Это позволяет защитной системе организма справиться с паразитами, оставшимися в недеформированных эритроцитах, прежде чем болезнь войдет в полную силу. Таким образом, в районах, где распространена малярия, гетерозиготность по гену серповидноклеточности дает человеку определенное преимущество. Хотя в гомозиготном состоянии этот ген обычно детален, для гетерозиготных индивидуумов он служит защитой от болезни, которая часто заканчивается смертью.
Болезнь Тея-Сакса, проявляющаяся в нарушении развития головного мозга и приводящая к смерти в возрасте около четырех лет, обусловлена наличием рецессивного летального гена. У индивидуумов, гомозиготных по этому гену, отсутствует фермент, ответственный за метаболические превращения определенного липида в клетках мозга. Без этого фермента липид накапливается, и клетки утрачивают способность нормально функционировать. Лечить эту болезнь пока не умеют. Известно, что среди еврейского населения Восточной Европы из каждых 30 человек один гетерозиготен по данному гену. Однако в США приблизительно одна треть всех случаев болезни Тея-Сакса приходится на долю детей из нееврейских семей.
Среди белого населения США чаще всего встречается летальный ген, обусловливающий кистозный фиброз - болезнь, при которой бронхиолы, а также протоки поджелудочной железы и печени заполняются густой слизью. Жертвы кистозного фиброза умирают обычно от инфекционных респираторных заболеваний в детском и юношеском возрасте, не доживая до 20 лет. Предполагают, что ген, ответственный за кистозный фиброз, кодирует один из ферментов цепи переноса электронов.
Серии множественных генов. Многие гены состоят из сотен нуклеотидов (а не тольков двух различных формах), так что мутации могут происходить во многих разных участках гена и порождать много различных новых его форм. Разумеется, каждый данный индивидуум несет в себе не более двух из таких разных форм, поскольку все гены представлены в организме парами, по одному в каждой из гомологичных хромосом.
Наиболее известным примером такого рода служит ген, определяющий группы крови системы АВО у человека. Этот ген встречается в трех главных формах: IА, IВ и i. Формы IА, и IВ кодируют два разных фермента, каждый из которых присоединяет к одному из белков на поверхности эритроцитов определенный сахар. Форма I не кодирует никакого фермента. У индивидуумов, несущих ген IА и ген IВ, синтезируются оба фермента, т.е. они кодоминантны. Ген i рецессивен по отношению к обоим этим генам. В таблице 2.1 представлены возможные генотипы и фенотипы для этой системы групп крови.
Таблица 2.1. Возможные генотипы для различных групп крови (фенотипов) в системе АВО у человека.
Группа крови (фенотип) |
Генотип |
Частота у населения США |
О I |
ii |
45% |
А II |
IАIА или IАi |
41% |
В III |
IВIВ или IВi |
10% |
АВ IV |
IАIВ |
4% |
Группы крови нашли интересное применение при установлении отцовства, когда, например, предполагаемому отцу предъявляется иск, или при подозрении, что детей перепутали в родильном доме. Достаточно нескольких капель крови, чтобы определить группу крови ребенка и его предполагаемых родителей, а затем на основании этих генетических данных решить, мог ли данный человек (или данная супружеская чета) произвести на свет ребенка с такой группой крови. Таким путем нельзя доказать, что именно этот мужчина или именно эта женщина является отцом или матерью данного ребенка, но часто можно доказать, что родителем этого ребенка данный человек быть не может. Если, например, человек с группой крови АВ имеет генотип IАIВ , то он не может быть отцом ребенка с группой крови О. Ребенок с группой крови О имеет генотип ii, и, стало быть, у его отца должен иметься хотя бы один ген i, который он мог бы передать своему ребенку (см. табл. 2.1). В то же время данный человек может быть отцом ребенка с группой крови А или В. Однако на свете есть много других людей, которые по своим группам крови могли бы быть отцом этого ребенка, а потому однозначно установить отцовство на основании групп крови нельзя.
Недавно для решения вопроса об отцовстве начали использовать белки, кодируемые другими сериями множественных генов, а именно белки, находящиеся на поверхности лейкоцитов. Существуют четыре разных гена, кодирующих эти белки, причем каждый представлен большим числом различных форм - от 9 до 16. Это обеспечивает столь большое разнообразие возможных генотипических комбинаций, что любой индивидуум (за исключением монозиготных близнецов) несет на клеточных мембранах своих лейкоцитов совершенно уникальные «химические отпечатки пальцев». Благодаря этому тест на лейкоцитарные белки можно использовать в тех случаях, когда тест на группы крови АВО при решении вопроса об отцовстве оказывается несостоятельным. Данная методика отличается очень высокой точностью, так что со времени ее введения в практику число отцов, которым приходится принимать на себя материальную ответственность за своих потомков, весьма ощутимо возросло.
Полигенные признаки. Наряду с отдельными генами, представленными множеством форм, существуют и полигенные признаки, т.е. признаки, контролируемые многими генами, находящимися в разных участках хромосомы, а иногда даже и в разных парах хромосом. У человека среди известных нам примеров этого рода можно назвать такие признаки, как рост, умственные способности, телосложение, а также цвет волос и цвет кожи.
Взаимодействие нескольких неаллельных генов с одинаковым действием называется полимерией, или однозначным действием генов.
Примером полимерии является наследование цвета кожи у человека. Этот признак определяется четырьмя генами, ответственными за выработку пигмента меланина. Чем больше активных генов, запускающих синтез меланина, имеется в клетке, тем темнее ее окраска. Эти гены локализованы в четырех парах разных хромосом. У людей с самой темной окраской кожи (негры) имеется восемь аллелей этих генов (поскольку клетки диплоидны): А1А1А2А2А3А3А4А4 (гены действуют одинаково, поэтому их обозначают одной и той же буквой). У человека с самой светлой кожей нет ни одного активного аллеля: а1а1а2а2а3а3а4а4. Дети таких двух людей получат четыре активных (доминантных) аллеля от одного из родителей и цвет их кожи будет промежуточным. Генотип детей будет: А1 а1А2 а2А3 а3А4 а4.
В зависимости от числа доминантных генов в генотипе может формироваться более светлый или более темный цвет кожи. Такой тип взаимодействия генов называется кумулятивной полимерией.
Полигенные признаки с трудом поддаются изучению, потому что непросто вычленить эффект каждого отдельного гена в данном фенотипическом признаке, отделив его от эффекта других генов. Влияние среды еще больше запутывает дело; скажем, в нашем примере с цветом кожи люди могут быть более или менее смуглыми в зависимости от интенсивности загара.
Генетика пола. Наследование, сцепленное с полом. В хромосомном наборе человека 22 пары хромосом представляют собой аутосомы – они не отличаются у мужчин и женщин. Лишь одна пара хромосом, называемых половыми, различна у мужчин и женщин (см. рис. 2.1). У женщин это две X-хромосомы, а у мужчин одна Х-хромосома и одна Y-хромосома. Половые хромосомы несут самые разнообразные гены, в том числе не имеющие отношения к первичным и вторичным половым признакам.
У человека развитие организма по мужскому типу определяет Y-хромосома. Если она отсутствует, развитие идет по женскому типу.
У женщин при образовании половых клеток в результате расхождения половых хромосом во всех яйцеклетках обязательно оказывается Х-хромосома. У мужчин в половине половых клеток оказывается Х-хромосома, а в другой половине – Y-хромосома. Пол будущего ребенка определяется в момент оплодотворения. Если в сперматозоиде будет Y-хромосома, то у возникшей в результате оплодотворения зиготы будут Х- и Y-хромосомы – этот набор обусловливает развитие мужчины. Если сперматозоид, оплодотворивший яйцеклетку, будет с Х-хромосомой, то в зиготе будут две Х-хромосомы и родится девочка.
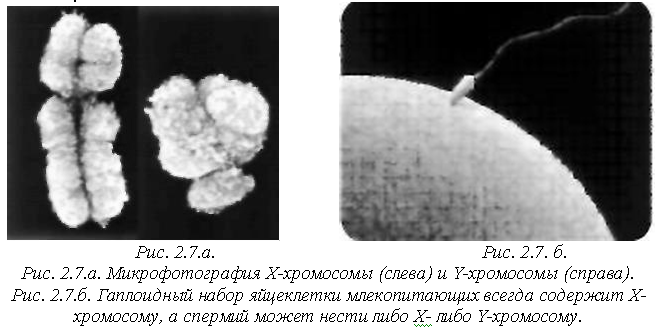
Х-хромосома по длине ДНК больше чем Y-хромосома приблизительно в три раза. На рисунке 2.7.а показано, как выглядят половые хромосомы человека.
А на рисунке 2.7.б показано, как выглядят клетки, содержащие эти хромосомы. Маленькая (сперматозоид) образована организмом генотипа XY, а большая – генотипа ХХ (яйцеклетка).
Н а
рисунке 2.8. видно как выглядит человек
после того, как он уже начал развиваться:
это игла, и на ее конце находится несколько
поделившихся клеток, полученных из
оплодотворенной яйцеклетки. Такой наш
размер на стадии 4-5-го деления.
а
рисунке 2.8. видно как выглядит человек
после того, как он уже начал развиваться:
это игла, и на ее конце находится несколько
поделившихся клеток, полученных из
оплодотворенной яйцеклетки. Такой наш
размер на стадии 4-5-го деления.
Данные, которыми мы располагаем, показывают, что у каждого зародыша имеются гены, необходимые для развития индивидуума как мужского, так и женского пола. Под действием гормонов из зародышей птиц и млекопитающих можно получать особей генетически «неправильного» пола. В нормальных условиях гормоны, вырабатываемые яичниками или семенниками, поддерживают у каждого индивидуума развитие «правильного» пола.
Как половые хромосомы определяют пол? На ранних стадиях развития у зародыша образуются зачаточные репродуктивные органы, не являющиеся ни мужскими, ни женскими. Что из них разовьется, зависит от половых хромосом. У человека в Y-хромосоме находится ген, под контролем которого определенные клетки продуцируют особый белок, локализующийся на клеточной поверхности. При наличии этого белка из репродуктивных органов зародыша развиваются семенники (мужские органы, которые со временем станут вырабатывать сперматозоиды). Семенники же в свою очередь вырабатывают гормоны, стимулирующие дифференцировку мужских половых путей.
С еменники
начинают дифференцироваться на 6-й
неделе зародышевого развития. Если
этого не произойдет, то из репродуктивных
органов на следующей неделе дифференцируются
яичники (женские органы, в которых
образуются яйцеклетки). В остальном
развитие женских половых путей происходит
автоматически, без каких-либо гормональных
сигналов со стороны яичников. Если
удалить яичники или семенники до того,
как произойдет дифференцировка половой
системы, то у зародыша разовьются женские
половые пути.
еменники
начинают дифференцироваться на 6-й
неделе зародышевого развития. Если
этого не произойдет, то из репродуктивных
органов на следующей неделе дифференцируются
яичники (женские органы, в которых
образуются яйцеклетки). В остальном
развитие женских половых путей происходит
автоматически, без каких-либо гормональных
сигналов со стороны яичников. Если
удалить яичники или семенники до того,
как произойдет дифференцировка половой
системы, то у зародыша разовьются женские
половые пути.
Итак, Y-хромосома несет по меньшей мере один ген, дающий зародышу первый «толчок» на пути его превращения в самца. Наличие Y-хромосомы обеспечивает мужскую природу, тогда как зародыши, несущие одни только X-хромосомы, превращаются в фенотипически женские особи. Из всех генов, участвующих в определении пола, только ген Y-хромосомы, «запускающий» дифференцировку семенников, локализован в половой хромосоме. Позднее вступают в действие многие другие гены, но все они находятся в неполовых хромосомах, в так называемых аутосомах.
На ранних стадиях зародышевого развития у самки млекопитающего функционируют обе Х-хромосомы. Позже, однако, во всех клетках (кроме тех, из которых разовьются яичники и яйцеклетки) одна из двух Х-хромосом инактивируется. Какая из Х-хромосом данной клетки утратит активность, определяется случайным образом; известно только, что у всех потомков этой клетки неактивной бывает одна и та же Х-хромосома. Такая неактивная Х-хромосома остается в конденсированной форме. Под микроскопом ее легко распознать в виде особой структуры, называемой тельцем Барра (рис 2.9).
Вследствие такой инактивации в любой клетке самки млекопитающего присутствует только одна активная Х-хромосома. Клетки самца млекопитающего также содержат по одной активной Х-хромосоме, поскольку в каждой из них имеется одна Х-хромосома и одна Y-хромосома. Присутствие или отсутствие тельца Барра позволяет распознавать мужские и женские клетки млекопитающих, например, для проверки пола в сомнительных случаях, на Олимпийских играх или других спортивных состязаниях. Тельце Барра служит также в некоторых экспериментах удобным «генетическим маркером».
Аномальное число половых хромосом возникает в тех случаях, когда нарушается расхождение хромосом в мейозе и в гаметы попадает на одну хромосому больше или, наоборот, меньше. Клетки, содержащие больше двух Х-хромосом, содержат и больше чем одно тельце Барра, потому что активна в них, как и всегда, только одна Х-хромосома.
Х- и Y-хромосомы сильно отличаются. Они гомологичны только в так называемых псевдоаутосомных зонах, которые присутствуют и в папиных и в маминых хромосомах, как и обычные аутосомы.
И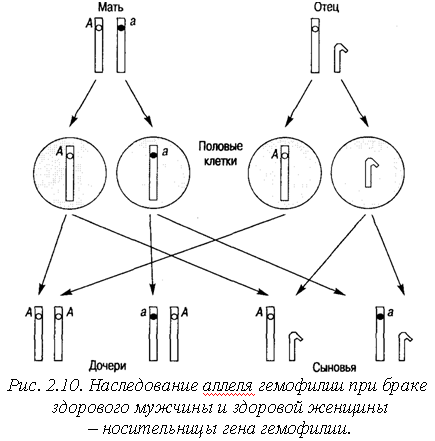 звестен
только один ген, определяющий пол; этот
ген находится в Y-хромосоме, а в Х-хромосоме
таких генов нет. Однако Х-хромосома
несет ряд других генов и, вероятно,
несколько генов локализуется также в
Y-хромосоме. У млекопитающих часть
Х-хромосомы гомологична части Y-хромосомы;
гены этих гомологичных участков ведут
себя так же, как и любые другие гены. Для
других участков Х-хромосомы соответствующих
участков в Y-хромосоме нет; находящиеся
здесь гены называют сцепленными с полом.
У самцов млекопитающих любой рецессивный
ген, локализующийся в одном из
негомологичных участков Х-хромосомы,
экспрессируется
в фенотипе,
потому что в Y-хромосоме нет гена, который
мог бы его маскировать. У самки такой
рецессивный ген проявляется в фенотипе
лишь в том случае, если он присутствует
в двух копиях. Вследствие этого
фенотипические признаки, определяемые
рецессивными сцепленными с полом генами,
встречаются у самцов млекопитающих
чаще, чем у самок. В этом заключается
одна из причин того, что смертность
среди самцов млекопитающих в любом
возрасте выше, чем среди самок: ведь
известно, что многие рецессивные гены
обладают вредным эффектом.
звестен
только один ген, определяющий пол; этот
ген находится в Y-хромосоме, а в Х-хромосоме
таких генов нет. Однако Х-хромосома
несет ряд других генов и, вероятно,
несколько генов локализуется также в
Y-хромосоме. У млекопитающих часть
Х-хромосомы гомологична части Y-хромосомы;
гены этих гомологичных участков ведут
себя так же, как и любые другие гены. Для
других участков Х-хромосомы соответствующих
участков в Y-хромосоме нет; находящиеся
здесь гены называют сцепленными с полом.
У самцов млекопитающих любой рецессивный
ген, локализующийся в одном из
негомологичных участков Х-хромосомы,
экспрессируется
в фенотипе,
потому что в Y-хромосоме нет гена, который
мог бы его маскировать. У самки такой
рецессивный ген проявляется в фенотипе
лишь в том случае, если он присутствует
в двух копиях. Вследствие этого
фенотипические признаки, определяемые
рецессивными сцепленными с полом генами,
встречаются у самцов млекопитающих
чаще, чем у самок. В этом заключается
одна из причин того, что смертность
среди самцов млекопитающих в любом
возрасте выше, чем среди самок: ведь
известно, что многие рецессивные гены
обладают вредным эффектом.
У человека хорошо известны такие рецессивные сцепленные с полом признаки, как цветовая слепота, гемофилия и один из видов мышечной дистрофии. Гемофилия также определяется рецессивным геном, находящимся в Х-хромосоме. У женщины, несущей такой ген, обычно имеется также в другой ее X-хромосоме доминантный нормальный ген, так что фенотипически она здорова - гемофилии у нее нет (рис. 2.10). Иначе обстоит дело у мужчины - у него нет второй Х-хромосомы, которая могла бы нести нормальный ген, у него ген гемофилии экспрессируется. В организме больного гемофилией почти не образуется того белка, который необходим для свертывания крови; такой человек может умереть - буквально истечь кровью от самого пустякового пореза. Гемофилию в наше время научились контролировать (но не излечивать!), вводя больным так называемый фактор свертывания, выделенный из крови здорового человека. Благодаря этому некоторые мужчины с гемофилией доживают теперь до такого возраста, что могут оставить потомство. Если такой мужчина вступит в брак с женщиной, гетерозиготной по гену гемофилии, то у них может родиться дочь с гемофилией. Дочь должна для этого унаследовать Х-хромосомы с геном гемофилии от обоих родителей. В прошлом больные гемофилией мужчины обычно умирали рано, не успев оставить потомство, поэтому мы и не знаем случаев, когда бы гемофилией болели женщины.
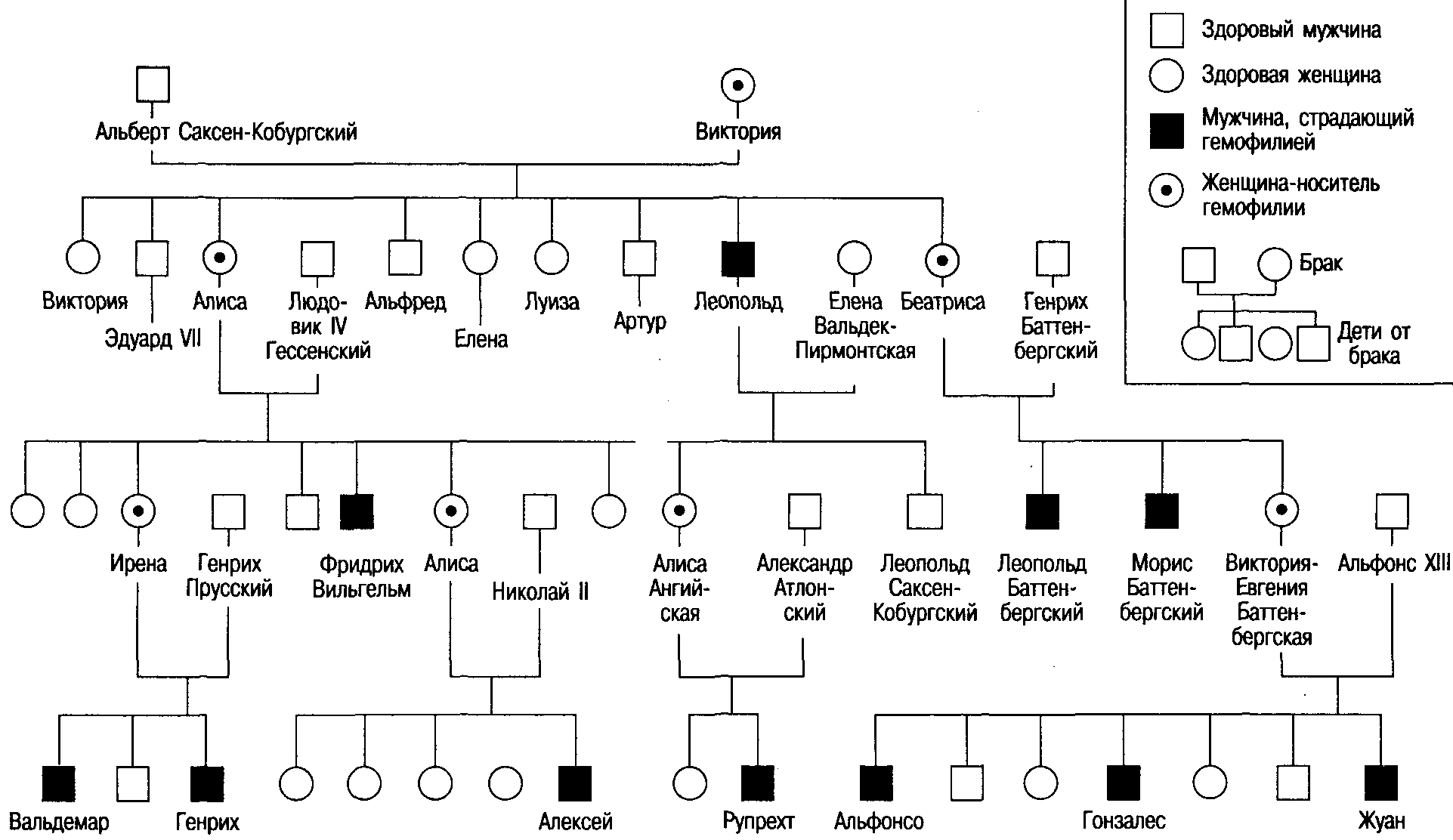
Рис. 2.11. Генеалогическое древо царствовавших семей Европы, иллюстрирующее наследование гена гемофилии, локализованного в Х-хромосоме.
На Х-хромосомах обозначены аллели рецессивного аллеля гемофилии а. Нормальный аллель доминирует – обозначен как А. Если в брак со здоровым мужчиной вступает здоровая женщина, являющаяся носительницей гена гемофилии, то с одинаковой вероятностью (в 25% случаев) может родиться здоровая дочь, здоровая дочь – носительница гена гемофилии, здоровый сын, больной сын. Таким образом, носительницами гена гемофилии являются женщины, а болеют ею мужчины. Однако если мужчина-гемофилик вступит в брак со здоровой женщиной, то все его сыновья будут обязательно здоровы (потому что они получат от отца Y-хромосому). Дочери будут тоже здоровыми, но обязательно будут носительницами гена гемофилии.
Известный всему миру пример носителя гемофилии - королева Виктория. Через ее страдавшего гемофилией сына Леопольда и через двух ее дочерей -Алису и Беатрису, являвшихся носительницами гемофилии, эта болезнь проникла в ряд королевских домов Европы, попала в Россию, Пруссию и Испанию. Прежде гемофилию называли «королевской болезнью», но ни один из ныне живущих монархов гена гемофилии не унаследовал (рис 2.11).Обратите внимание, что у Леопольда, больного гемофилией, был здоровый сын, а здоровая дочь унаследовала ген гемофилии.
Теоретически возможна гемофилия и у женщины, но такая вероятность очень невысока, так как для этого необходимо вступление в брак больного-гемофилика с женщиной-носительницей гена гемофилии (и даже в этом случае вероятность рождения больной девочки будет только 0,25). Из-за низкой частоты встречаемости гена гемофилии и того, что больные гемофилией часто умирают до брачного возраста, такие случаи практически не отмечаются.
Если гены локализованы в Y-хромосоме, то они должны передаваться только от отцов к сыновьям. В качестве примера такого гена обычно упоминают ген, вызывающий появление пучка волос на внешнем крае уха. Недавно сообщалось об обнаружении гена-маркера на Y-хромосоме, который сцеплен с геном, ответственным за мужскую гипертонию. Если на Y-хромосоме обнаруживается ген-маркер, то у мужчин систолическое давление выше в среднем на 10 мм ртутного столба. Часть ядерных генов также наследуется только однородительски. Это гены Y-хромосомы, которая наследуется только от отца к сыну. Те гены в Y-хромосоме, которые не имеют пары в Х-хромосоме, наследуются однородительски – от отца к сыну.
Факторы, влияющие на экспрессию генов. От наследования, сцепленного с полом, надо отличать наследование, ограниченное полом. Все гены, которыми обладает данный организм, определяют только его генетический потенциал, т.е. только то, каким он может быть. Иное дело - каким он окажется в действительности. Зародышевое развитие зависит от взаимодействия всех генов во время их экспрессии, иными словами в то время, когда они обеспечивают - или не обеспечивают образование тех или иных полипептидов и белков. Важнейшую роль в развитии играют также факторы среды. В последние два десятилетия мы несколько раз были свидетелями поистине страшных случаев, когда под влиянием лекарственных препаратов, принимаемых беременными женщинами, нарушалось нормальное развитие плода и рождались уроды, или когда дети этих женщин в раннем возрасте заболевали раком (вследствие утраты их организмом контроля над клеточным делением).
Роль половых гормонов заключается главным образом в воздействии на репродуктивную систему и связанные с нею органы, однако эти гормоны могут также влиять на ряд других признаков организма. Гены, степень экспрессии которых определяется уровнем половых гормонов, называются генами, зависимыми от пола. (Обычно, хотя и не всегда, такие гены локализуются в аутосомах.) Бык может, например, нести гены, определяющие высокую молочность, но молока он давать не будет, потому что у него слишком низок уровень женских гормонов. Эти гены делают его, однако, ценным производителем для молочного стада. Точно так же, и самцы и самки обладают генетическим потенциалом, необходимым для образования органов, свойственных противоположному полу, но у них в процессе развития образуются органы, характерные для их собственного пола, потому что уровень соответствующих гормонов у них выше. У самок и у самцов имеются также гормоны, свойственные противоположному полу, но содержание их гораздо ниже.
Ген, определяющий облысение, типичное для мужчин, локализован в аутосоме, но его экспрессия зависит от мужских половых гормонов. У мужчин этот ген ведет себя как доминантный ген из-за присутствия мужских половых гормонов; у женщин же он ведет себя как рецессивный ген, так что женщина лысеет в том случае, если у нее имеется две дозы этого гена.
Пол влияет у человека и на такой признак, как подагра. При подагре в тканях, главным образом в суставах (чаще всего в области большого пальца стопы), отлагаются соли мочевой кислоты, причиняющие человеку мучительную боль. Ген, ответственный за это заболевание, экспрессируется в присутствии мужских половых гормонов гораздо сильнее, нежели в присутствии женских. В литературе викторианской эпохи подагра фигурирует главным образом как одна из причин частых приступов гнева у капризных старых джентльменов. Считалось, что для облегчения страданий больным надо воздерживаться от жирной и острой пищи и не пить красного вина. Эти ограничения, однако, лишь еще больше портили характер жертв подагры. К счастью, в наше время подагру умеют лечить.
Половые гормоны далеко не единственные факторы, влияющие на фенотипическую экспрессию генов. Многие признаки, контролируемые в основном только одной парой генов, в какой-то мере зависят от влияния продуктов других генов, называемых генами-модификаторами. Долгое время считалось, что цвет глаз у человека определяется одной парой генов, причем карий цвет доминирует над голубым. Теперь мы знаем, что в определении цвета глаз участвуют по меньшей мере также и две пары генов-модификаторов и что у голубоглазых родителей может родиться кареглазый ребенок, хотя это и случается крайне редко.
В разном возрасте организм вырабатывает разные гормоны, так что и возраст тоже играет роль в экспрессии генов. Достаточно вспомнить, например, многие изменения, сопровождающие половое созревание: у мальчиков - «ломку» голоса и рост семенников; у девочек - увеличение молочных желез и появление характерных жировых отложений, придающих женской фигуре свойственную ей округлость; и, наконец, у обоих полов - рост волос в подмышечных впадинах и на лобке.
На экспрессию генов влияют также факторы внешней среды, а именно пища, свет и температура. Так, плохо питающиеся люди бывают обычно ниже ростом, чем допускают их гены. Сейчас во многих странах молодые люди обогнали в росте своих отцов именно потому, что они с детства питались лучше, чем их родители.
Свет - один из факторов, влияющих на экспрессию генов. Человек, подвергающийся в течение некоторого времени действию солнечных лучей, становится от этого более смуглым (некоторые, впрочем, только краснеют!).
Вопросы для закрепления:
В чем заключается революционный характер открытия Г. Менделя?
Как можно доказать, что наследственные факторы дискретны и парны?
Какие выводы были сделаны Г. Менделем на основании расщепления при моногибридном скрещивании?
Что такое доминантный и рецессивный признаки?
Что такое дигибридное скрещивание и какой вид при этом имеет расщепление?
Как выглядит решетка Пеннета для дигибридного скрещивания?
Как можно представить себе дигибридное скрещивание у человека?
Что такое рекомбинация и при образовании каких клеток она происходит?
Что такое хромосома?
Что такое диплоидный и гаплоидный наборы хромосом?
В каких клетках человека имеется гаплоидный набор хромосом?
Какие виды изменчивости вы знаете?
Что такое норма (диапазон) реакции?
Какие типы наследования выделяют?
Каки гены называют зависимыми от пола?
Что такое фенотипическая экспрессия мутантных генов?
Как происходит наследование аллеля гемофилии?
Какие факторы, влияют на экспрессию генов?
Почему один ген является доминантным, а другой рецессивным?
Кого называют носителями, а кого - больными?
Приведите примеры летальных генов.
Приведите примеры серии множественных генов.
Что означает однозначное действие генов?
Литература: [1, 6, 9, 10, 11, 12, 13,17,18].