

Тема 3. Основы популяционной генетики
Каждый живой организм, в том числе и человеческий, обладает целым набором присущих ему свойств. Некоторые из них являются общими для всех представителей данного вида (видоспецифические особенности). Например, каждый представитель вида Homo sapiens отличается от представителей других видов способностью к прямохождению, отсутствием волосяного покрова на большей части тела, высокоразвитым интеллектом и способностью к речевой коммуникации. Это все видоспецифические особенности. Каждый индивид, помимо видоспецифических характеристик, обладает целым набором индивидуальных, присущих только ему, качеств - это индивидуально-специфические особенности. Все эти особенности, присущие данному индивиду - и видоспецифические, и индивидуальные - в генетике принято называть признаками Виды, населяющие Землю, образуют сообщества, то есть пространственно-временные объединения. Причиной образования сообществ является одинаковая приспособленность особей к определенным экологическим условиям. Например, в пустынях не встречаются животные, биология которых требует высокой влажности климата. Одной из разновидностей сообществ является популяция. Поскольку генетика поведения работает в основном на уровне популяций, рассмотрим, как определяется это понятие в генетике.
►Популяция - совокупность свободно скрещивающихся особей одного вида, длительно существующей на определенной территории (части ареала), и относительно обособленной от других совокупностей того же вида. Важнейший признак популяции – это относительно свободное скрещивание. Если возникают какие-то изоляционные барьеры, препятствующие свободному скрещиванию, то возникают новые популяции.
►Идеальной популяцией в генетике считается группа скрещивающихся особей, живущих на одной территории. Если вероятности скрещиваний между особями не зависят от каких-либо дополнительных обстоятельств (возраста особей, половых предпочтений и т.п.), то мы имеем дело со случайно скрещивающейся популяцией. Иными словами, в такой популяции любая особь (или индивид) одного пола имеет равные шансы скрещивания (заключения брака) с любой особью (индивидом) другого пола. Случайно скрещивающуюся популяцию иногда называют панмиксной (термин «панмиксия» означает свободное случайное скрещивание; идеальная панмиксия возможна лишь в очень больших популяциях, не подвергающихся давлению отбора, мутаций и других факторов). Многие теоретические модели в психогенетике (и вообще в популяционной генетике) основаны на предположении о наличии в популяции свободного случайного скрещивания.
Принцип свободного и случайного скрещивания в популяциях может нарушаться, если образование пар по какому-либо признаку происходит не случайно. Например, в человеческих популяциях имеется тенденция к неслучайному подбору супружеских пар по росту. В генетике такой неслучайный подбор пар носит название «ассортативность». Например, в отношении роста говорят, что в популяции существует ассортативность по росту. Наличие или отсутствие ассортативности можно проверить, оценивая сходство супругов: если корреляция по данному признаку близка к нулю, то говорят, что ассортативность отсутствует; если корреляция отличается от нуля, это означает наличие ассортативности.
Ассортативность браков, т. е. наличие определенных предпочтений при выборе супругов может сказываться на фенотипической изменчивости. Она приводит к отклонению от законов менделевских популяций (одно из требований, которым должна удовлетворять такая популяция, - свободное скрещивание в пределах популяции). Чаще всего наблюдается положительная ассортативность, когда при вступлении в брак наблюдается положительная корреляция по какому-либо признаку между супругами.
О тмечается
положительная ассортативность браков
по коэффициенту интеллекта (рис. 3.1).
Сильнее всего она выражена для крайних
значений признака - лица с высоким
коэффициентом интеллекта предпочитают
вступать в брак с лицами с высоким
коэффициентом (коэффициент корреляции
0,3-0,4). При низком значении коэффициента
интеллекта тоже выбирают себе подобных.
В последнем случае коэффициент корреляции
особенно высок (0,68). При положительной
ассортативности в популяции происходит
увеличение генетической дисперсии, так
как увеличивается разнообразие генотипов.
тмечается
положительная ассортативность браков
по коэффициенту интеллекта (рис. 3.1).
Сильнее всего она выражена для крайних
значений признака - лица с высоким
коэффициентом интеллекта предпочитают
вступать в брак с лицами с высоким
коэффициентом (коэффициент корреляции
0,3-0,4). При низком значении коэффициента
интеллекта тоже выбирают себе подобных.
В последнем случае коэффициент корреляции
особенно высок (0,68). При положительной
ассортативности в популяции происходит
увеличение генетической дисперсии, так
как увеличивается разнообразие генотипов.
Существует положительная ассортативность по таким признакам, как рост, музыкальные способности, социально-экономическое положение, склонность к нейротизму. Отмечена ассортативность браков даже по некоторым параметрам ЭЭГ: очевидно, характеристики ЭЭГ влияют на какие-то особенности поведения, значимые для выбора партнера. В некоторых случаях встречается отрицательная ассортативность: например, рыжеволосые редко вступают в брак друг с другом. По ряду параметров личности ассортативность не обнаружена, например по темпераменту.
Популяции не являются статичными образованиями. В них постоянно происходят процессы миграции, имеются колебания рождаемости и смертности и другие изменения. Эти процессы приводят к колебаниям численности популяции и изменению частот встречаемости различных признаков.
Поскольку человек относится к одному из видов живых организмов, для него также характерно образование популяций. Помимо биологических причин, на формирование человеческих популяций влияют и различные социальные факторы. Человеческие популяции могут иметь различную численность. Термин «популяция» можно применить к населению таких мегаполисов, как Москва или Токио (в которых ежедневно происходят процессы миграции, приводящие к обновлению и изменению численности за счет притока или оттока населения), и к небольшим сообществам или племенам, населяющим замкнутые территории, например, в горной местности. Замкнутые популяции носят название изолятов. Изоляты в человеческих популяциях могут встречаться не только в географически изолированных местностях, но и внутри крупных популяций.
У человека, например, помимо территориальной изоляции, достаточно изолированные популяции могут возникать на основе социальных, этнических или религиозных барьеров. Поскольку между популяциями не происходит свободный обмен генами, то они могут существенно различаться по генетическим характеристикам. Для того чтобы описывать генетические свойства популяции, вводится понятие генофонда – совокупности генов, встречающихся в данной популяции. Помимо генофонда важна также частота встречаемости гена, или частота встречаемости аллеля.
С реди
представителей любой популяции
наблюдается определенная изменчивость,
то есть разнообразие признаков у ее
членов. Изменчивость человеческой
популяции легко заметить, если посмотреть
на толпу людей. Наряду с видоспецифическими
признаками каждый индивид обладает
уникальным набором признаков, присущих
только ему. В результате в толпе людей
практически невозможно обнаружить двух
одинаковых индивидов. Индивиды различаются
не только по внешнему строению - по цвету
волос и глаз, росту и телосложению, но
и по особенностям поведения - манере
говорить, походке, жестикуляции и т.п.
реди
представителей любой популяции
наблюдается определенная изменчивость,
то есть разнообразие признаков у ее
членов. Изменчивость человеческой
популяции легко заметить, если посмотреть
на толпу людей. Наряду с видоспецифическими
признаками каждый индивид обладает
уникальным набором признаков, присущих
только ему. В результате в толпе людей
практически невозможно обнаружить двух
одинаковых индивидов. Индивиды различаются
не только по внешнему строению - по цвету
волос и глаз, росту и телосложению, но
и по особенностям поведения - манере
говорить, походке, жестикуляции и т.п.
В зависимости от типа изменчивости в популяциях можно выделить признаки двоякого рода. Одни из них характеризуются дискретным, то есть прерывистым рядом изменчивости. Эти признаки встречаются в ограниченном числе вариантов, различия между индивидами четко выражены, и промежуточные формы отсутствуют. Признаки такого рода относят к категории качественных (рисунок 3.3). Их еще называют качественными . Внешние условия относительно мало или почти совсем не влияют на их проявление. К таким признакам относятся, например, группы крови у человека. Носители разных вариантов качественных признаков в популяции могут встречаться с разной частотой: можно вспомнить, что люди с резус-отрицательным фактором крови встречаются гораздо реже, чем с резус-положительным. Кроме того, частота встречаемости вариантов одного и того же признака в разных популяциях обычно различается.
На рисунке 3.2 представлен пример распределения частот встречаемости наиболее известных групп крови системы АВ0 в популяциях русских и коренных жителей Австралии. Можно заметить, что частоты встречаемости всех четырех групп крови в этих популяциях отличаются. Среди признаков человека, так или иначе связанных с поведением (в широком смысле), признаков качественных, имеющих дискретный характер изменчивости, почти нет. Из тех, что не связаны с грубой патологией, наиболее известны два, имеющие отношение к сенсорной чувствительности. Это чувствительность к вкусу фенилтиомочевины (одни люди ощущают вкус этого вещества, а другие - нет) и цветовая слепота - дальтонизм (некоторые люди не способны различать отдельные цвета спектра). Большинство же известных на сегодняшний день простых качественных признаков человека связано с биохимическими факторами крови или наследственными аномалиями, не имеющими значительных проявлений в поведении.
Ч асто
качественные полиморфизмы, которым
сопутствуют определенные особенности
поведения, связаны с различными
наследственными аномалиями, приводящими
к дегенеративным изменениям в центральной
нервной системе, сопровождающимся
нарушениями поведения. Примерами таких
полиморфизмов, затрагивающих поведение,
являются фенилкетонурия - нарушение
метаболизма,
приводящее к тяжелой умственной
отсталости, и хорея Гентингтона -
дегенеративное заболевание нервных
клеток, приводящее к непроизвольным
движениям, изменениям личности и
постепенно нарастающему слабоумию.
Основная
масса поведенческих характеристик
человека относится ко второй категории
признаков, обладающих непрерывной,
или количественной,
изменчивостью - континуалъностъю
проявлений. Подобную изменчивость мы
наблюдаем в таких морфологических
признаках, как рост, вес, цвет волос и
кожи, и в таких поведенческих признаках,
как интеллект, черты темперамента и
т.д. Все значения каждого из этих признаков
можно разместить на некоторой непрерывной
шкале. Очень высокие и очень низкие
значения признака, как правило, встречаются
в популяциях реже, чем средние. Частота
встречаемости тех или иных значений
признака может быть подсчитана и
представлена в виде распределения,
которое является совокупностью значений
количественного признака и соответствующих
им частот. Частотное распределение по
признаку, проявляющему непрерывную
изменчивость, примерно соответствует
кривой нормального распределения (рис.
3.4).
асто
качественные полиморфизмы, которым
сопутствуют определенные особенности
поведения, связаны с различными
наследственными аномалиями, приводящими
к дегенеративным изменениям в центральной
нервной системе, сопровождающимся
нарушениями поведения. Примерами таких
полиморфизмов, затрагивающих поведение,
являются фенилкетонурия - нарушение
метаболизма,
приводящее к тяжелой умственной
отсталости, и хорея Гентингтона -
дегенеративное заболевание нервных
клеток, приводящее к непроизвольным
движениям, изменениям личности и
постепенно нарастающему слабоумию.
Основная
масса поведенческих характеристик
человека относится ко второй категории
признаков, обладающих непрерывной,
или количественной,
изменчивостью - континуалъностъю
проявлений. Подобную изменчивость мы
наблюдаем в таких морфологических
признаках, как рост, вес, цвет волос и
кожи, и в таких поведенческих признаках,
как интеллект, черты темперамента и
т.д. Все значения каждого из этих признаков
можно разместить на некоторой непрерывной
шкале. Очень высокие и очень низкие
значения признака, как правило, встречаются
в популяциях реже, чем средние. Частота
встречаемости тех или иных значений
признака может быть подсчитана и
представлена в виде распределения,
которое является совокупностью значений
количественного признака и соответствующих
им частот. Частотное распределение по
признаку, проявляющему непрерывную
изменчивость, примерно соответствует
кривой нормального распределения (рис.
3.4).
Большинство людей попадает в среднюю часть распределения, а на его краях, представляющих крайние степени выраженности признака, оказывается лишь малая часть популяции.
Часто при оценке количественных признаков мы пользуемся качественными категориями, такими, как «высокий-низкий», «сильный-слабый», «темный-светлый» и т.п. Вспомним известный пример из психофизиологии. Людей часто делят на имеющих сильный и слабый тип нервной системы, однако такое деление условно. В эти группы попадают только люди с краев распределения, тогда как сам параметр силы-слабости измеряется по непрерывной шкале, и в популяции встречаются все значения, начиная от крайней слабости нервной системы и заканчивая крайними значениями силы. Точно так же весьма условно деление людей на экстравертов и интровертов. Любое типологическое деление не описывает полной картины изменчивости по континуальному признаку.
С уществует
еще одна категория признаков, занимающая
как бы промежуточное положение между
качественными полиморфизмами и
количественными, континуальными
признаками. Речь идет о признаках
с пороговым эффектом.
Внешне эти признаки напоминают
качественные полиморфизмы, то есть
отличаются дискретным характером
проявления. Организм либо несет данный
признак, либо нет. Чаще всего признаками
с пороговым эффектом являются различные
заболевания, например, сахарный
диабет,
бронхиальная астма или шизофрения,
а также различные врожденные аномалии
развития - анэнцефалия, spina bifida (аномалии
центральной нервной системы), заячья
губа, волчья пасть и др. Для этих болезней
и пороков развития наблюдаются четкие
альтернативные распределения, подобные
распределениям по качественным
полиморфизмам: индивид либо страдает
данным заболеванием, либо нет. Однако
тип наследования при этих заболеваниях
оказывается ближе к типу наследования
количественных признаков. В этом случае
можно сказать, что порог является
условной границей в нормальном
распределении признака, разделяющей,
с одной стороны, пораженных, и с другой
- непораженных, но предрасположенных к
заболеванию индивидов. При некоторых
заболеваниях граница между нормой и
патологией прослеживается довольно
четко, особенно при врожденных аномалиях
развития, тогда как при других она
размыта и весьма условна (например,
граница между умственной отсталостью
и нормальным интеллектом). Таким образом,
признаки с пороговым эффектом скорее
могут быть отнесены к категории
количественных признаков человека.
Соответственно, для них будут справедливы
те же закономерности наследования, что
и - для обычных континуальных
признаков
(рис. 3.5).
уществует
еще одна категория признаков, занимающая
как бы промежуточное положение между
качественными полиморфизмами и
количественными, континуальными
признаками. Речь идет о признаках
с пороговым эффектом.
Внешне эти признаки напоминают
качественные полиморфизмы, то есть
отличаются дискретным характером
проявления. Организм либо несет данный
признак, либо нет. Чаще всего признаками
с пороговым эффектом являются различные
заболевания, например, сахарный
диабет,
бронхиальная астма или шизофрения,
а также различные врожденные аномалии
развития - анэнцефалия, spina bifida (аномалии
центральной нервной системы), заячья
губа, волчья пасть и др. Для этих болезней
и пороков развития наблюдаются четкие
альтернативные распределения, подобные
распределениям по качественным
полиморфизмам: индивид либо страдает
данным заболеванием, либо нет. Однако
тип наследования при этих заболеваниях
оказывается ближе к типу наследования
количественных признаков. В этом случае
можно сказать, что порог является
условной границей в нормальном
распределении признака, разделяющей,
с одной стороны, пораженных, и с другой
- непораженных, но предрасположенных к
заболеванию индивидов. При некоторых
заболеваниях граница между нормой и
патологией прослеживается довольно
четко, особенно при врожденных аномалиях
развития, тогда как при других она
размыта и весьма условна (например,
граница между умственной отсталостью
и нормальным интеллектом). Таким образом,
признаки с пороговым эффектом скорее
могут быть отнесены к категории
количественных признаков человека.
Соответственно, для них будут справедливы
те же закономерности наследования, что
и - для обычных континуальных
признаков
(рис. 3.5).
Когда мы анализировали моно- и дигибридное скрещивание и рисовали решетку Пеннета, мы имели дело с отдельной семьей и потомством от единичного скрещивания. Теперь нашим объектом будет популяция.
В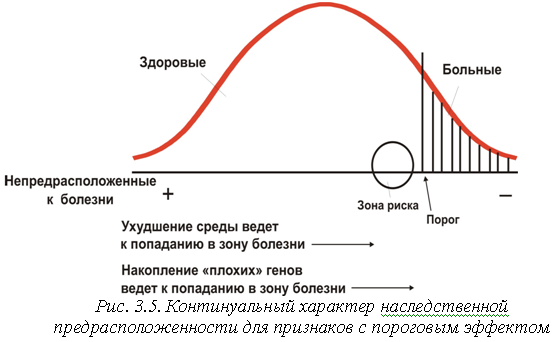 качестве удобной модели расщепления
по одной паре аллелей
может служить наследование групп крови
системы MN. Группа крови этой системы
определяется двумя аллелями М и N.
Гомозиготы
ММ имеют группу крови М, гомозиготы NN
имеют группу крови N, а гетерозиготы
MN имеют группу крови MN.
качестве удобной модели расщепления
по одной паре аллелей
может служить наследование групп крови
системы MN. Группа крови этой системы
определяется двумя аллелями М и N.
Гомозиготы
ММ имеют группу крови М, гомозиготы NN
имеют группу крови N, а гетерозиготы
MN имеют группу крови MN.
Рассмотрим замкнутую человеческую популяцию, в которой имеется какое-то количество аллелей М и какое-то количество аллелей N. В принципе, частота встречаемости аллелей М и N в популяции может меняться от 100% М, когда вся популяция представлена только гомозиготами ММ, до 100% N, когда вся популяция представлена только гомозиготами NN. Если аллели встречаются с одинаковой частотой, то частота встречаемости каждого из них составит 50%, или 0,5. Предположим, что в нашей популяции представлены не только гомозиготы одного типа, а все три типа сочетаний аллелей, и частота встречаемости каждого аллеля составляет 0,5. Понятно, что в такой популяции с равной вероятностью будут производиться гаметы, несущие аллель М и аллель N, т.е. частоты встречаемости этих гамет также будут равны 0,5. Если браки в этой популяции происходят случайно, то мы можем воспользоваться решеткой Пеннета и изобразить частоту образования гомозигот и гетерозигот в следующем поколении (вероятности перемножаются).
|
Женские гаметы |
|
Мужские гаметы |
0,5 М |
0,5 N |
0,5 М 0,5 N |
0,25 ММ 0,25 MN |
0,25 MN 0,25 NN |
Мы видим, что частоты встречаемости гомозигот (ММ и NN) составляют по 0,25, тогда как частота встречаемости гетерозигот (MN) в два раза выше - 0,5. Частоты же встречаемости каждого аллеля (М и N) будут по-прежнему одинаковыми - по 0,5. Соответственно в следующем поколении этой популяции (F2), при сохранении случайности браков, будут опять получены те же соотношения.
В реальных популяциях, как правило, наблюдаются самые различные частоты встречаемости аллелей, к тому же между аллелями могут существовать отношения доминантности и рецессивности, и гетерозиготы по внешней выраженности признака могут совпадать с доминантным типом гомозигот, т.е. частоты встречаемости самого признака будут отличаться от частот встречаемости гомозигот и гетерозигот.
Разберем еще один пример, связанный со вкусовой чувствительностью. Когда мы говорили о дискретной изменчивости, мы упоминали о существовании двух типов людей, имеющих разную чувствительность к вкусу фенилтиомочевины (ФТМ). Эта чувствительность зависит от одного гена, представленного парой аллелей. Доминантный аллель Т определяет чувствительность к вкусу ФТМ, а рецессивный аллель t - отсутствие чувствительности. Таким образом, гомозиготы tt не ощущают горького вкуса ФТМ, тогда как гомозиготы ТТ и гетерозиготы Tt его ощущают. Опять представим себе изолированную человеческую популяцию, в которой браки осуществляются случайно, а частоты встречаемости аллелей Т и t составляют 0,6 и 0,4 соответственно. Естественно, что частоты гамет, несущих данные аллели, будут такими же, и мы опять можем изобразить с помощью решетки Пеннета частоту различных сочетаний аллелей в поколении F1.
|
Женские гаметы |
|
Мужские гаметы |
0,6 Т |
0,4 t |
0,6 Т 0,4 t |
0,36 ТТ 0,24 tT |
0,24 Tt 0,16 tt |
Таким образом, в поколении F1 частоты гомозигот и гетерозигот будут следующими:
ТТ = 0,62 = 0,36;
Tt = 2.0,4.0,6 = 0,48;
tt = 0,42 = 0,16.
Заметим, что в сумме это составляет единицу. Если рассмотреть частоту встречаемости самого признака, то окажется, что в данной популяции число людей, не ощущающих вкус ФТМ, составит 16%.
Т = 0,36 + 1/2.0,48 = 0,6;
t = 0,16 + 1/2.0,48 = 0,4;
Отсюда ясно, что частоты аллелей при замкнутости популяции и случайности браков сохранились и в следующем поколении.
Все это может быть записано в виде буквенных выражений. Если частоты аллелей обозначить буквами р и q, то мы можем записать, что в популяции р + q = 1. Решетку Пеннета для аллелей Т и t мы можем представить так:
|
Женские гаметы |
|
Мужские гаметы |
pТ |
qt |
рТ qt |
р2ТТ pqTt |
pqTt q2tt |
Таким образом, мы можем записать, что сумма частот встречаемости различных зигот равна: р2 + 2pq + q2 = (р + q)2 .
Эта формула отражает распределение гомозигот и гетерозигот в популяции. Она была впервые предложена в 1908 г. двумя исследователями - математиком Г. Харди и врачом В. Вайнбергом, независимо друг от друга.Таким образом, закон Харди–Вайнберга позволяет подсчитывать частоты аллелей в популяции. Рецессивные аллели проявляются в фенотипе, только если они оказываются в гомозиготном состоянии. Гетерозиготы фенотипически либо не отличаются от доминантных гомозигот, либо их можно идентифицировать с помощью специальных методов, подчас весьма сложных.
Пусть в популяции представлены два аллеля А и а, с частотой встречаемости соответственно p и q. Тогда p + q=1.
Несложные подсчеты показывают, что в условиях свободного скрещивания относительные частоты генотипов АА, Аа, аа будут составлять соответственно р2, 2pq, q2. Суммарная частота, естественно, равна единице:
р2+ 2pq + q2=1.
► Закон Харди-Вайнберга гласит, что в условиях идеальной популяиии частоты генов и генотипов остаются постоянными от поколения к поколению.
Для выполнения закона Харди-Вайнберга требуются несколько условий.
Например, необходима случайность скрещивания в популяции - одинаковая вероятность скрещивания между всеми особями, входящими в состав популяции. Нарушения этого условия у человека могут быть связаны с кровнородственными браками. В этом случае в популяции повышается количество гомозигот.
Еще одна причина нарушения закона Харди-Вайнберга – это ассортативность браков, которая связана с неслучайностью выбора брачного партнера. Например, обнаружена определенная корреляция между супругами по коэффициенту интеллекта. Ассортативность быть может быть положительной или отрицательной и соответственно повышать изменчивость в популяции или уменьшать ее. Отметим, что ассортативность влияет не на частоты аллелей, а на частоты гомо- и гетерозигот.
Эти положения в естественных условиях в той или иной степени нарушаются. Однако в целом их влияние не так сильно выражено и в человеческих популяциях соотношения Харди-Вайнберга, как правило, выполняются.
В каждом поколении частота каждого аллеля данного гена и частота каждого генотипа по этому гену сохраняется постоянной. В этом случае и частоты фенотипов постоянны.
Произведем расчеты для рецессивной мутации, вызывающей заболевание фенилкетонурию. Заболевание встречается у 1 человека на 10 тыс. Таким образом, частота встречаемости гомозигот q2 (генотип аа) равна 0,0001. Частота рецессивного аллеля q определяется путем извлечения квадратного корня и равна 0,01.Частота доминантного аллеля будет равна: p =1- q = 1- 0,01 = 0,99.
Отсюда легко определить частоту встречаемости гетерозигот А:
2pq = 2 х 0,99 х 0,01 = 0,0198 ~ 0,02, т. е. она составляет приблизительно 2%. Получается, что 1 человек из 50 является носителем гена фенилкетонурии. Эти данные показывают, какое большое число рецессивных генов остается в скрытом состоянии.
При близкородственном скрещивании (инбридинге) частота гомозиготных генотипов увеличивается по сравнению с соотношениями закона Харди-Вайнберга. В результате этого вредные рецессивные мутации, определяющие заболевания, чаще оказываются в гомозиготном состоянии и проявляются в фенотипе. Среди потомства от кровнородственных браков с большей вероятностью встречаются наследственные заболевания и врожденные уродства.
Показано, что с увеличением степени инбридинга снижаются показатели умственного развития и школьная успеваемость. При увеличении коэффициента инбридинга на 10% коэффициент интеллекта снижается на 6 баллов (по шкале Векслера для детей). Коэффициент инбридинга в случае брака двоюродных сибсов равен 1/16, для троюродных сибсов – 1/32. Например, частота генетически наследования заболевания фенилкетонурия при неродственных браках составляет 1:15000, а при родственных – 1:7000; альбинизма – 1:40000 и 1:3000 соответственно.
В связи с повышением мобильности населения в развитых странах и разрушением изолированных популяций наблюдается снижение коэффициента инбридинга в течение всего XX в. На это также повлияло снижение рождаемости и уменьшение числа двоюродных сибсов.
При отдаленном скрещивании можно наблюдать появление гибридов с повышенной жизнеспособностью в первом поколении. Это явление получило название гетерозиса. Причиной гетерозиса является перевод вредных рецессивных мутаций в гетерозиготное состояние, при котором они не проявляются в фенотипе.
Цель психогенетики - определить роль факторов наследственности и среды в формировании индивидуальных различий по психологическим и психофизиологическим признакам. Необходимо оценить изменчивость признака в фенотипе для данной популяции и попытаться дать ответ па вопрос об относительном вкладе в эту изменчивость генетических и средовых факторов.
Популяционный подход к оценке наследуемости особенностей поведения не позволяет описывать процессы взаимодействия генотипа и среды в индивидуальном развитии. Когда в результате психогенетических исследований, проведенных, скажем, на близнецах или на приемных детях, признак относят к наследуемому, это не значит, что он наследственно детерминирован в общепринятом смысле этого слова. На первый взгляд, звучит парадоксально. Психогенетические исследования ведутся в основном на популяционном уровне. Когда на основании коррелирующего поведения у родственников популяционные генетики делают вывод о наследуемости признака, это не означает, что индивидуальное развитие данного поведения обусловлено исключительно генетическими причинами.
Высокая наследуемость свидетельствует лишь о том, что разнообразие индивидов в популяции в значительной степени связано с генотипическими различиями между ними. Имеется в виду, что процент индивидов, обладающих данным признаком в популяции потомков, может быть предсказан, исходя из знаний о родительской популяции. Однако значение показателя наследуемости ничего не говорит о последовательности событий в индивидуальном развитии признака и о том, какой конечный фенотип будет результатом развития конкретного индивида. В этом смысле признак с высокой оценкой наследуемости не является детерминированным генотипом, хотя такие интерпретации часто встречаются даже в публикациях специалистов. Это совсем разные вещи - разделить источники вариативности в популяции на генетические и средовые или искать генетические и средовые причины, лежащие в основе онтогенетического формирования конкретных фенотипов.
Результат развития - фенотип - зависит от совместного действия генов и среды. Гены и признаки связаны сложной сетью путей развития. Все индивидуальные различия, которыми занимаются дифференциальные психологи и психогенетики, являются результатом обстоятельств развития конкретных индивидов в конкретных средах. Часто индивиды, воспитанные в явно различающихся средах, имеют много общего. И наоборот, сиблинги, воспитывающиеся в одной семье, казалось бы при сходных обстоятельствах, за счет тонких различий в условиях воспитания и развития реально будут испытывать весьма различные воздействия как физической, так и социальной среды. Это справедливо даже для генетически идентичных МЗ близнецов. Таким образом, процесс взаимодействия со средой сложен и неоднозначен. Отметим также, что психологи и другие исследователи часто пользуются термином «взаимодействие» в статистическом смысле, когда исследуется взаимодействие отдельных факторов в продуцировании какого-либо измеряемого эффекта. Подчеркнем, что статистическое взаимодействие факторов и взаимодействие генов и среды в индивидуальном развитии - это совершенно разные вещи. Их не следует путать.
Фенотипические различия между людьми объясняются по крайней мере двумя причинами. Во-первых, люди отличаются друг от друга своими генотипами. Это приводит к возникновению генетически обусловленной изменчивости. Во-вторых, каждый человек развивается в особенных средовых условиях. Это приводит к возникновению средовой изменчивости.
Фенотипическое выражение признака в популяции определяется его средним значением и дисперсией. Для оценки изменчивости удобно применять дисперсию, т. е. меру изменчивости признака:
г де
VP
- фенотипическая
дисперсия признака, Xi,
- значение признака у i-го
члена популяции, X
- среднее
значение признака в популяции, N
- число
индивидов в изучаемой популяции.
де
VP
- фенотипическая
дисперсия признака, Xi,
- значение признака у i-го
члена популяции, X
- среднее
значение признака в популяции, N
- число
индивидов в изучаемой популяции.
В генетике для обозначения дисперсии используется латинская буква V (от английского variance). Значками VP VG и VE обозначаются соответственнофенотипическая (P = phenotype), генотипическая (G = genotype) и средовая (Е = environment) дисперсии.
Часть наблюдаемой изменчивости признака в популяции связана с тем, что имеются разные генотипы, т. е. отличия между индивидами связаны с отличиями в генах - генотипическая дисперсия (VG).
Другая часть изменчивости связана с различиями в средовых воздействиях - средовая дисперсия (VE).
При изучении вклада генетических и средовых влияний на формирование различий в поведении человека фенотипическое разнообразие (фенотипическая дисперсия VP )разлагают на два компонента: генетический и средовой. Часть наблюдаемой изменчивости признака в популяции связана с тем, что имеются разные генотипы, т. е. отличия между индивидами связаны с отличиями в генах - генотипическая дисперсия (VG). Другая часть изменчивости связана с различиями в средовых воздействиях - средовая дисперсия (VE). В качестве меры разнообразия выступает дисперсия (варианса):
г де
Vp
- фенотипическая дисперсия; VA
- аддитивная компонента; VD
- компонента, отражающая отклонение
фенотипа от ожидаемого значения в
аддитивной модели; VEB
- межсемейная компонента, показывающая
различия между членами
разных семей; VEW
- «случайная» средовая
компонента, складывающаяся из различий
между членами одной семьи; VGE
- компонента дисперсии, возникающая в
результате взаимодействия генотипа со
средой (например, социабельность
экстравертов и интровертов повышается,
если первые воспитываются в больших
семьях, а вторые - в малочисленных); CovGE
- часть дисперсии, обусловленная
неслучайным распределением генотипов
в разных средах (экстраверты, как правило,
находятся в более разнообразной
социальной
среде, чем интроверты); VM
компонента,
отражающая варьирование тест-ретес-товых
измерений (ошибку измерения).
де
Vp
- фенотипическая дисперсия; VA
- аддитивная компонента; VD
- компонента, отражающая отклонение
фенотипа от ожидаемого значения в
аддитивной модели; VEB
- межсемейная компонента, показывающая
различия между членами
разных семей; VEW
- «случайная» средовая
компонента, складывающаяся из различий
между членами одной семьи; VGE
- компонента дисперсии, возникающая в
результате взаимодействия генотипа со
средой (например, социабельность
экстравертов и интровертов повышается,
если первые воспитываются в больших
семьях, а вторые - в малочисленных); CovGE
- часть дисперсии, обусловленная
неслучайным распределением генотипов
в разных средах (экстраверты, как правило,
находятся в более разнообразной
социальной
среде, чем интроверты); VM
компонента,
отражающая варьирование тест-ретес-товых
измерений (ошибку измерения).
Рассмотрим, как общая популяционная изменчивость признаков складывается из генетической и средовой изменчивости (компьютерная модель количественной изменчивости в Л.С. Куравского). Допустим, мы измерили некий количественный признак у 1000 индивидов в какой-либо популяции и построили соответствующее частотное распределение. Будем считать, что наш признак определяется одним геном, который может существовать в трех различных формах, т.е. имеет три аллеля. Обозначим их буквами А, В и С. Тогда в популяции могут встретиться три типа гомозигот - АА, ВВ, СС - и три типа гетерозигот - АВ, ВС и АС. Пусть люди с генотипом АА имеют самые низкие значения признака, а люди с генотипом СС - самые высокие. Генотип ВВ занимает промежуточное положение. Эффекты доминирования отсутствуют, и гетерозиготы оказываются промежуточными между гомозиготами.
Мы знаем, что на конкретную величину признака оказывают влияние не только гены, но и средовые условия, например, характер питания или родительской заботы в детстве. Распределение фенотипов для каждого генотипа будет характеризоваться определенным средним значением и разбросом вокруг среднего, или дисперсией. Величина дисперсии будет определяться тем, насколько данный генотип чувствителен к средовым влияниям. Чем выше чувствительность генотипа к среде, тем большее разнообразие фенотипов мы получим и соответственно тем большей дисперсией будет характеризоваться данный генотип (см. предыдущий параграф). Понятно, что внутри группы особей с одинаковым генотипом дисперсия будет определяться только средовыми факторами и поэтому будет чисто средовой. Например, для представителей генотипа АА: VP(AA) = VE(AA)
Предположим, мы смогли разделить людей, обладающих конкретными генотипами, на соответствующие группы, то есть мы разбили всю популяцию на шесть групп в соответствии с генотипом каждого человека. Обычно отдельные генотипы представлены в популяции неравномерно, поэтому можно ожидать, что и обладатели наших вымышленных генотипов встречаются с определенной частотой.
Таким образом, можно измерить: среднее значение признака для каждого генотипа, частоту его встречаемости в популяции, величину разброса индивидуальных значений вокруг среднего, то есть дисперсию VE(AA), VE(BB) и т.д.
Представим, что проведены все измерения, получены статистические оценки и они занесены в таблицу (количественные значения признака были выбраны таким образом, чтобы они примерно соответствовали реальному количественному признаку человека, каковым является рост в сантиметрах, поэтому можно представить е, что мы имеем дело с формированием популяционной изменчивости по росту, хотя, конечно же, генетическая детерминация реального роста отличается от схемы, приведенной в данной модели).
Одновременно мы построили частотные распределения для каждого конкретного генотипа и общепопуляционное распределение (рис. 3.6).На графике мы видим шесть субраспределений, каждое из которых описывает один из генотипов в популяции. Распределение каждого генотипа представлено пропорционально его встречаемости в популяции. Наибольшей представленностью на графике характеризуется генотип АВ, поскольку этот генотип более всего распространен в популяции, а наименьшей - СС. Одни генотипы (АА, АВ) характеризуются небольшими средними значениями признака, другие (ВС, СС) - большими. Каждому из генотипов соответствует определенное разнообразие фенотипов (дисперсия), поскольку обладатели этого генотипа росли в разных средах. Если генотип очень чувствителен к среде (например, BB и АС), ему будет соответствовать большая фенотипическая дисперсия, а если малочувствителен (например, СС), то выраженность признака у всех обладателей этого генотипа будет близка к генотипической средней. Форма же общепопуляционного распределения является следствием сложения всех частных распределений для каждого из генотипов.
Теперь обратимся к таблице. В одной из колонок выписаны значения средних величин признака для каждого генотипа. Наибольший интерес представляет последняя колонка, в которой представлены дисперсии. Мы видим, что дисперсии генотипов значительно различаются вследствие их разной чувствительности к средовым влияниям:
Генотип |
Размер выборки |
Частота в популяции |
Среднее значение признака |
Дисперсия |
АА |
100 |
од |
150 |
20,4 |
АВ |
400 |
0,4 |
165 |
50,1 |
вв |
300 |
0,3 |
170 |
149,7 |
АС |
150 |
0,15 |
175 |
142,6 |
ВС |
30 |
0,03 |
190 |
105,2 |
ОС |
20 |
0,02 |
200 |
5,8 |
П ри
этом дисперсия внутри каждого генотипа
целиком определяется влияниями среды.
Общая же изменчивость в популяции и
характеризующая ее величина
общепопуляционной дисперсии складывается
из двух составляющих. Одна из них - это
различия между генотипами, выражающиеся
в различиях между средними значениями
признака у обладателей отдельных
генотипов (АА, АВ и т.д.), а другая - это
различия внутри каждого из генотипов,
которые определяются только средой. В
нашем примере генетическая составляющая
дисперсии возникает при участии только
шести генотипов, то есть генетическая
дисперсия возникла в результате различий
между средними значениями признака
имеющихся у нас шести генотипов. Если
на графике убрать кривые распределений,
а оставить только столбцы, соответствующие
частоте встречаемости средних значений
признака этих шести генотипов, то мы
будем иметь шесть таких столбцов,
представляющих гистограмму распределения
средних значений признака всех генотипов
нашей популяции. В реальных популяциях,
где имеется множество различных
генотипов, такая гистограмма будет
состоять не из шести столбцов, а из сотен
и тысяч, а распределение будет
соответствовать нормальному. Средовая
составляющая общепопуляционной дисперсии
складывается из отдельных средовых
дисперсий внутри каждого генотипа.
Таким образом, общепопуляционная
дисперсия состоит из средовой дисперсии
внутри отдельных генотипов и генетической
дисперсии, возникающей за счет различий
между генотипами, что можно выразить
формулой VP = VG
+ VE.
ри
этом дисперсия внутри каждого генотипа
целиком определяется влияниями среды.
Общая же изменчивость в популяции и
характеризующая ее величина
общепопуляционной дисперсии складывается
из двух составляющих. Одна из них - это
различия между генотипами, выражающиеся
в различиях между средними значениями
признака у обладателей отдельных
генотипов (АА, АВ и т.д.), а другая - это
различия внутри каждого из генотипов,
которые определяются только средой. В
нашем примере генетическая составляющая
дисперсии возникает при участии только
шести генотипов, то есть генетическая
дисперсия возникла в результате различий
между средними значениями признака
имеющихся у нас шести генотипов. Если
на графике убрать кривые распределений,
а оставить только столбцы, соответствующие
частоте встречаемости средних значений
признака этих шести генотипов, то мы
будем иметь шесть таких столбцов,
представляющих гистограмму распределения
средних значений признака всех генотипов
нашей популяции. В реальных популяциях,
где имеется множество различных
генотипов, такая гистограмма будет
состоять не из шести столбцов, а из сотен
и тысяч, а распределение будет
соответствовать нормальному. Средовая
составляющая общепопуляционной дисперсии
складывается из отдельных средовых
дисперсий внутри каждого генотипа.
Таким образом, общепопуляционная
дисперсия состоит из средовой дисперсии
внутри отдельных генотипов и генетической
дисперсии, возникающей за счет различий
между генотипами, что можно выразить
формулой VP = VG
+ VE.
В рассмотренном примере количественная оценка общей дисперсии оказалась равной 168,7, а усредненная оценка средовой дисперсии - 91,7, то есть почти вполовину меньше. Если из величины общей дисперсии вычесть величину средовой дисперсии, мы получим оценку генетической дисперсии. Таким образом, в нашем примере генетическая дисперсия равна:168,7-91,7=77,0.
Это означает, что генетическая дисперсия обусловливает чуть меньше половины всех индивидуальных различий в популяции
Таким образом, основными компонентами фенотипической дисперсии признака являются генетическая и средовая дисперсии.
►Генотипическая дисперсия признака - это изменчивость признака, связанная с изменчивостью генотипов; средовая дисперсия признака - дисперсия, связанная с дисперсией средовых воздействий:
VP=VG+VE.
В количественной генетике долю генетической составляющей в общей фенотипической дисперсии признака принято называть наследуемостью в широком смысле слова, или просто наследуемостью. Наследуемость обычно обозначают символом h2. Наследуемость можно подсчитать, если величину генетической дисперсии разделить на величину общей дисперсии:
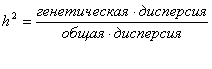 или
или
![]()
Если использовать данные рассмотренного нами примера, то
![]()
В данном выражении h2 - это коэффициент наследуемости в широком смысле, поскольку VG включает все компоненты генотипической дисперсии. Иногда говорят о коэффициенте генетической детерминации. Коэффициент наследуемости может принимать значения от 0 до 1.
Таким образом, доля генетической составляющей в фенотипической дисперсии в нашем примере составляет 46%. Это означает, что изменчивость (вариативность) нашего признака в популяции на 46% складывается за счет вариативности генотипов и на 54% - за счет вариативности сред. Показатель наследуемости - это генетическая составляющая дисперсии. Поэтому при символе наследуемости всегда стоит значок квадрата, указывающий на то, что показатель наследуемости есть компонент дисперсии.
Наследуемость является одним из основных показателей, которыми оперирует генетика поведения. Если наследуемость отлична от нуля, это означает, что в основе изменчивости признака лежат не только средовые, но и генетические причины. Когда задается вопрос, лежат ли в основе индивидуальных психологических различий наследственные причины, для ответа необходимо получить данные о влиянии генов на фенотипическую изменчивость. Величина коэффициента наследуемости и позволяет оценить меру влияния генотипа на изменчивость признака.
Вся история генетики поведения до некоторого времени представляла собой бесконечные попытки обосновать существование наследственных причин межиндивидуальных различий в поведении. После создания теории наследственности количественных признаков и разработки математического аппарата для анализа генетических и средовых дисперсий практически в каждой работе по генетике поведения человека приводятся данные о наследуемости изучавшейся психологической характеристики. Когда в печати стали появляться сообщения о том, что наследуемость умственных способностей составляет 70-80%, это привлекло внимание не только ученых, но и общества в целом, и оказало в некоторых странах влияние на социальную политику в отношении умственно неполноценных людей. Если коэффициент интеллекта имеет высокую наследуемость, стоит ли тратить силы и средства на специальное обучение индивидов со сниженным интеллектом? И в целом ряде случаев из-за некомпетентного подхода к пониманию наследуемости общество предпринимало ошибочные шаги в решении отдельных социальных проблем.
Негативные социальные последствия психогенетических исследований интеллекта явились в том числе результатом очень распространенного заблуждения, связанного с неверным толкованием понятия «наследуемость». К сожалению, очень часто научное понятие наследуемости смешивается с обыденным пониманием наследуемого как чего-то рокового, не подверженного изменению под влиянием средовых условий. Возможно, это обусловлено самой терминологией, поскольку слово «наследуемость» сходно по звучанию со словом «наследственность». Остановимся более подробно на том, какое же содержание вкладывается в научное понятие наследуемости.
Уточним составные компоненты фенотипической дисперсии:
VP=VG + VE.
Дисперсии можно складывать при условии, что между ними отсутствует корреляция; если же она имеет место, добавляются поправочные компоненты:
VP=VG+ VE + 2CovGE + VGE+ju + VM,
где CovGE- ковариация (корреляция) генотип - среда, VCE - взаимодействие «генотип-среда», ju- ассортативность браков, VM - дисперсия измерений (ошибка измерений, случайные флуктуации при повторных измерениях).
Рассмотрим подробно вклад каждого компонента в общую изменчивость фенотипа.
Генотипический компонент (VG состоит из нескольких компонентов, отражающих разные типы взаимодействия генов:
простой вариант суммирования эффектов генов (аддитивный тип взаимодействия);
эпистатическое взаимодействие (эпистаз), когда одни гены влияют на экспрессию других генов;
доминантный тип взаимодействия генов.
Основной вклад в генотипическую дисперсию вносит аддитивный тип взаимодействия (VA), поэтому можно учитывать только его.
Мутации увеличивают генотипическую дисперсию, селекция в результате естественного отбора или генетический дрейф - уменьшают.
Средовой компонент VE состоит из двух составных частей:
VE=VS+VNS.
Разделенная среда (shared) -VS представляет такие влияния среды, как стиль родительского воспитания, доходы семьи, уровень интеллектуальной стимуляции в доме, т. е. одинаково воздействующие на родственников. Таким образом, разделенная среда может быть потенциальным источником их сходства. Другое ее название - общая среда.
Неразделенный компонент, или индивидуальная среда, - VNS, VS, - это воздействия, которые неодинаковы у родственников, воспитываемых вместе (происшествия, отношения со сверстниками, различное обращение со стороны родителей). Эти средовые воздействия могут быть источником индивидуальных различий.
Исследования поведения человека показывают, что основной источник средовых влияний на психологические свойства человека - это индивидуальные средовые воздействия, то, что относится к понятию «неразделенная среда».
Взаимодействие «генотип-среда» - (GxE), VGE - сказывается в том, что среда оказывает разное действие на особи с разным генотипом. Одно и то же средовое воздействие влияет на разные генотипы по-разному, вызывает разную реакцию, приводит к появлению разных свойств, признаков, как количественных, так и качественных. Например, исследования алкоголизма или девиантного поведения часто указывают на наличие генетически обусловленной уязвимости по отношению к обычным воздействиям внешней среды. Наличие определенного генотипа здесь связано с особой реакцией на средовые воздействия, которые не оказывают влияния на другие генотипы. В таких случаях речь идет не просто об изменчивости генотипов, определяющей вариации признака в фенотипе, а о свойстве генотипа определять изменчивость, которая возникает под влиянием внешней среды.
Корреляция «генотип-среда» (rGE), или «ковариация» CovCE. В этом случае генотип определяет часть средовых воздействий.
При пассивной корреляции (rСЕ) речь идет о «наследовании» среды в соответствии с наследственными задатками. Ребенок получает от родителей не только определенный генотип, но и среду, в которой воспитывается и которая может влиять на развитие его наследственных склонностей.
В случае реактивной корреляции (rGE) окружающая среда определенным образом реагирует на свойства, связанные с генетическими особенностями человека. Иногда эту корреляцию называют эвокативной (evocative), подчеркивая, что генотип вызывает реакцию среды. Например, родители могут заметить математические или музыкальные способности ребенка и определить его в соответствующую школу. Если проявление этих способностей было связано с генетическими предпосылками, то мы имеем право говорить в этом случае о реактивной корреляции (rСЕ). Родители по-разному ведут себя с капризным и раздражительным ребенком или с послушным и веселым.
Корреляция может повысить изменчивость, когда воздействие среды усиливает развитие свойств, связанных с наличием генетических задатков, которые и вызвали реакцию среды. Могут наблюдаться случаи отрицательной генотип-средовой корреляции, в результате которой происходит уменьшение изменчивости по данному признаку. Например, в школе зачастую основное внимание уделяется отстающим ученикам, которых стараются подтянуть до нормы. Отрицательная генотип-средовая корреляция часто сопровождает приведение личностных характеристик к биосоциальной норме в обществе.
Активная корреляция (rGE) связана с тем, что человек с какими-то особенностями генотипа может выбирать для себя или создавать соответствующую среду. Лонгитюдные исследования часто обнаруживают зависимость между поведением детей и характеристиками их среды во взрослом возрасте. Если данные особенности детского поведения генетически обусловлены, то это результат активной корреляции. Если выбор среды происходит по причинам, не связанным с особенностями генотипа, то это не является генотип-средовой корреляцией.
Чисто аддитивный характер взаимодействия генов встречается достаточно редко, чаще всего картина является более сложной. Для некоторых генов могут иметь место уже известные нам эффекты доминирования. При этом степень доминирования может отличаться для разных генов. В известных нам опытах Г. Менделя один из аллелей полностью подавлял другой, то есть имело место полное доминирование. Однако могут встречаться и пары аллелей, для которых доминантность и рецессивность не проявляются в полной мере. Это случаи неполного доминирования, или кодоминантности. В случае неполного доминирования гетерозиготы обладают промежуточным фенотипом между родительскими формами.
Поскольку генотипическая дисперсия практически определяется только аддитивной компонентой, употребляют понятие наследуемости в узком смысле:
![]()
Коэффициент наследуемости в этом случае отражает вклад аддитивных генетических факторов в наблюдаемую фенотипическую изменчивость.
Когда мы говорим о низком коэффициенте наследуемости, то это не означает, что отсутствует генетическая детерминация признака. Подавляющее большинство признаков детерминированы генетически. Речь идет о том, что в наблюдаемом индивидуальном разнообразии по данному признаку генотип не «виноват». Наследуемость может быть вообще равна нулю, если гены, участвующие в формировании данного признака, будут одинаковы у всех особей в популяции. Изменчивость признака, связанная с генетической изменчивостью, будет равна нулю, и коэффициент наследуемости соответственно тоже будет равен нулю. Наследственность и наследуемость - это разные понятия.
Н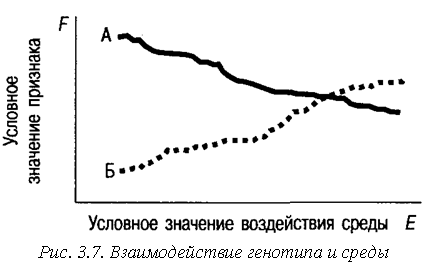 аследуемость
- это характеристика не признака вообще.
Это характеристика признака в данной
популяции, при данной совокупности
условий среды. В другой популяции, при
иных воздействиях среды, значения
наследуемости могут быть другими.
аследуемость
- это характеристика не признака вообще.
Это характеристика признака в данной
популяции, при данной совокупности
условий среды. В другой популяции, при
иных воздействиях среды, значения
наследуемости могут быть другими.
Если признак обладает высоким коэффициентом наследуемости, то это не означает, что он нечувствителен к средовым воздействиям. Может показаться, что если наследуемость равна 90%, то бесполезно менять средовые воздействия, ничего не изменится, все равно на признак уже не повлиять. По величине генетической дисперсии в данных условиях нельзя предсказать, как она будет меняться под влиянием изменения среды. Теоретически возможны ситуации, когда генетическая дисперсия признака может исчезнуть под влиянием среды. Представим, что в популяции имеются два разных генотипа А и Б, по-разному реагирующие на изменение среды (рис. 3.3). Тогда возможно такое состояние среды, при котором средние значения признака для каждого генотипа совпадут (точка пересечения). В этой точке для данной популяции генотипическая дисперсия исчезнет и останется только средовая.
При уменьшении разнообразия средовых воздействий наследуемость будет возрастать, поскольку в этих условиях индивидуальные отличия в большей мере будут связаны с генетическими факторами. В принципе коэффициент наследуемости может быть равен единице. Это может также наблюдаться для жестко детерминированных признаков, практически не зависящих от средовых воздействий. Например, такие признаки, как цвет глаз, группа крови, практически не зависят от средовых воздействий, и вся наблюдаемая изменчивость определяется генотипической изменчивостью.
Для любого генотипа существует диапазон сред, в котором он может проявить себя «максимально»; среду, одинаково благоприятную для всех генотипов, найти нельзя. Дело не в обогащенности сред, а в их качественном разнообразии. Сред должно быть много, чтобы у каждого генотипа была возможность найти «свою» среду и реализоваться. Однообразная среда, какой бы обогащенной она ни была, будет благоприятствовать развитию только определенных, а не всех генотипов.
Если считать, что генотипы равномерно распределены по средам (т.е. оказываются в определенной среде совершенно случайно), то компонент 2covGE становится равным 0, и тогда мы опять имеем ту формулу фенотипической структуры популяции, которая рассматривалась нами как исходная:VP = VG + VE. В реальных популяциях генотип-средовая ковариация является обычным явлением и ее вклад в вариативность признаков, как правило, не равен нулю. Действительно, трудно себе представить, чтобы индивид совершенно случайно попадал в ту или иную среду. С рождением каждого ребенка популяция пополняется еще одним генотипом, но ребенок обычно сразу же попадает в среду своей собственной семьи, к своим собственным родителям, бабушкам, дедушкам, братьям и сестрам, с которыми у него есть общие гены. Если при игре в рулетку брошенный шарик случайно оказывается в ячейке, то только что родившийся ребенок занимает «ячейку» вполне определенную: в своей семье, своем городе, своей стране. Таким образом, его среда не может быть совершенно случайной.
Р ассмотрим
следующую схему (рис. 3.8). Предположим,
в попу-ляции существуют две семьи (А и
В). Супруги А, оба, обладают «плохими»
генами в отношении какого-то количественного
признака с аддитивным типом наследования
(допустим, это интеллект), т.е. у них
преобладают гены-ослабители. В то же
время в другой семье (В) супруги имеют
«хорошие» гены (преобладают гены-усилители).
В результате рекомбинаций при образовании
гамет дети супругов А и супругов В могут
получить различные сочетания генов, и,
в принципе, среди них могут оказаться
обладатели сходных сочетаний (зона
перекрытия на рисунке), однако все же
вероятность того, что дети супругов А
окажутся обладателями «худших» генов
достаточно велика, и, наоборот, дети
супругов В скорее всего получат «лучшие»
гены. На схеме генотипы детей обозначены
точками а и b на соответствующей оси
(внизу). Теперь представим себе, в какой
среде окажутся дети в этих семьях. Скорее
всего в силу более высоких умственных
способностей родители В обеспечат
своему ребенку лучшую среду для развития,
чем родители А. В семье В среда будет
обогащенной (больше книг, развивающих
игрушек, дополнительных занятий, речевое
общение более высокого уровня и т.д.), а
в семье А - обедненной, поскольку у
родителей, вероятно, будет меньше
возможностей создать подобную среду.
ассмотрим
следующую схему (рис. 3.8). Предположим,
в попу-ляции существуют две семьи (А и
В). Супруги А, оба, обладают «плохими»
генами в отношении какого-то количественного
признака с аддитивным типом наследования
(допустим, это интеллект), т.е. у них
преобладают гены-ослабители. В то же
время в другой семье (В) супруги имеют
«хорошие» гены (преобладают гены-усилители).
В результате рекомбинаций при образовании
гамет дети супругов А и супругов В могут
получить различные сочетания генов, и,
в принципе, среди них могут оказаться
обладатели сходных сочетаний (зона
перекрытия на рисунке), однако все же
вероятность того, что дети супругов А
окажутся обладателями «худших» генов
достаточно велика, и, наоборот, дети
супругов В скорее всего получат «лучшие»
гены. На схеме генотипы детей обозначены
точками а и b на соответствующей оси
(внизу). Теперь представим себе, в какой
среде окажутся дети в этих семьях. Скорее
всего в силу более высоких умственных
способностей родители В обеспечат
своему ребенку лучшую среду для развития,
чем родители А. В семье В среда будет
обогащенной (больше книг, развивающих
игрушек, дополнительных занятий, речевое
общение более высокого уровня и т.д.), а
в семье А - обедненной, поскольку у
родителей, вероятно, будет меньше
возможностей создать подобную среду.
Следовательно, среда семьи В будет способствовать максимальному развитию исходных задатков своего ребенка, и его фенотип будет постоянно улучшаться. В то же время среда семьи А не создаст возможностей для развития фенотипа ребенка. В результате наследственно заложенные задатки не проявятся в полной степени. Таким образом, мы имеем ситуацию, в которой совершенно пассивно, просто по факту своего рождения, обладатель лучшего генотипа унаследовал и лучшую среду, а обладатель худшего генотипа унаследовал худшую среду.
Здесь мы имеем дело с явлением, которое в психогенетике называется пассивной положительной ковариацией генотипа и среды. Ковариация (корреляция) всегда положительна, если два параметра изменяются (ковариируют) в одном направлении. Например, чем больше рост, тем больше длина ступни человека. Это пример положительной ковариации. В рассмотренном нами случае генотип и среда также ковариируют положительно, поскольку лучшему генотипу соответствует лучшая среда и наоборот. Пассивной ковариация генотипа и среды называется потому, что ни родители, ни дети не прикладывают специальных усилий для создания среды: дети просто «наследуют» среду так же, как и гены. Это называется культурным наследованием.
На той же схеме (рис.) стрелками показано, что среда как бы «тянет» фенотипы (Pa) и (Pb) в разные стороны. Понятно, что в результате та разница, которая имелась между детьми при рождении (диапазон фенотипических различий - D0), будет усугубляться (диапазон фенотипических различий - Di). Если такая картина наблюдается не только в одной семье, а типична для популяции вообще, то результатом генотип-средовой ковариации будет увеличение фенотипической дисперсии, поскольку разброс между фенотипами в результате такой ковариации, как мы видим, возрастает.
Теперь рассмотрим еще одну подобную схему (рис. 3.9). Предположим, что в семье В родился второй ребенок (b2), которому «не повезло», и он получил худшее сочетание генов. Родители обнаружили, что второй ребенок не столь одарен от природы, как первый, и направили все усилия на его развитие, забросив первого, который «и сам пробьется». Тем самым они создали худшему генотипу лучшие условия среды. Это пример отрицательной генотип-средовой ковариации, поскольку переменные варьируют в различных направлениях (хуже генотип - лучше среда и, наоборот, лучше генотип - хуже среда). В результате фенотипические различия между детьми начнут уменьшаться (конечный диапазон различий Di меньше начального - D0). Таким образом, отрицательная генотип-средовая ковариация должна уменьшать популяционную дисперсию, но это в том случае, если рассмотренный пример представляет собой типичное явление. Напоминаем, что ковариация является величиной статистической и может быть обнаружена только на значительных выборках. Если же подобные усилия родителей наблюдаются лишь в одной семье, а в других происходит нечто иное, то никакой отрицательной ковариации генотипа и среды мы не получим.
Т олько
что рассмотренная ситуация является
примером отрицательной реактивной
ковариации генотипа и среды. Реактивной
обычно называется ковариация, возникающая
в результате специальных усилий
социального окружения ребенка в ответ
на какие-либо его специфические
особенности, связанные с наследственной
конституцией. Например, дети, проявляющие
признаки природной одаренности в спорте,
музыке, изобразительной деятельности,
привлекают внимание взрослых (родителей,
педагогов, воспитателей), и те стараются
создать условия для дальнейшего развития
природных задатков: помещают детей в
специализированные школы, кружки по
интересам и т.п. В результате «хороший»
генотип получает «хорошую» среду, т.е.
наблюдается положительная ковариация.
олько
что рассмотренная ситуация является
примером отрицательной реактивной
ковариации генотипа и среды. Реактивной
обычно называется ковариация, возникающая
в результате специальных усилий
социального окружения ребенка в ответ
на какие-либо его специфические
особенности, связанные с наследственной
конституцией. Например, дети, проявляющие
признаки природной одаренности в спорте,
музыке, изобразительной деятельности,
привлекают внимание взрослых (родителей,
педагогов, воспитателей), и те стараются
создать условия для дальнейшего развития
природных задатков: помещают детей в
специализированные школы, кружки по
интересам и т.п. В результате «хороший»
генотип получает «хорошую» среду, т.е.
наблюдается положительная ковариация.
Реактивная ковариация может быть и отрицательной. Она может возникать, например, в том случае, когда наследственной отягощенности или какому-либо неблагополучию ребенка, обусловленному наследственными причинами, противопоставляются компенсирующие усилия родителей, врачей, педагогов, заставляющие работать среду против генотипа. Однако это скорее исключение, чем правило. Большинство детей с проблемами развития оказываются в среде, которая только усугубляет их неблагополучие. Например, большие трудности часто возникают у детей с гиперактивностью и нарушениями внимания, нарушениями эмоционально-волевой сферы, «трудным» темпераментом. Все это можно назвать синдромом «трудного» ребенка. К сожалению, такие «особые» дети чаще вызывают отрицательные реакции окружающих: их больше наказывают, унижают и обижают, они реже бывают успешными в различных видах деятельности. Получается, что их природное (чаще всего оно и наследственное) неблагополучие влечет за собой и неблагоприятную среду. Ковариация генотипа и среды в этом случае будет положительной, поскольку и генотип, и среда действуют в одном направлении («плохой» генотип влечет за собой «плохую» среду). Безусловно, это приводит к отрицательным последствиям для фенотипа, но генотип-средовая ковариация в данном случае положительная.
Усилия практических психологов должны быть направлены на преодоление существующей тенденции. Необходимо помогать родителям, воспитателям и педагогам создавать такие условия среды, которые будут улучшать фенотип ребенка, имеющего ту или иную отягощенность. Практический психолог, участвуя в отборе детей в специализированные школы, консультируя родителей, участвуя в организации среды в детских учреждениях, должен помнить, что от его усилий зависит, произойдут ли положительные сдвиги в развитии ребенка. Поэтому при «плохом» генотипе следует создавать отрицательную ковариацию генотипа и среды, а при «хорошем» - положительную.
Мы рассмотрели два типа ковариации - пассивную и реактивную - и обнаружили, что она может быть как положительной, так и отрицательной. Если вернуться к формуле: VP = VG + VE +2covGE,то становится понятным, что положительная ковариация должна увеличивать фенотипическую дисперсию в популяции, а отрицательная - уменьшать.
В психогенетике, помимо пассивной и реактивной форм ковариации генотипа и среды, выделяют еще и активную ковариацию. Она характерна для более старших возрастов, начиная с подросткового, когда индивид самостоятельно (активно) выбирает ту среду, которая ему более подходит. Часто это происходит против воли родителей и других взрослых. Типичным примером активной генотип-средовой ковариации является судьба самого Ф. Гальтона, который по настоянию отца долгое время готовился к карьере врача, но после смерти отца забросил медицину и хотя и продолжал изучать природу человека, но уже в совершенно ином аспекте. Таким образом, получается, что не столько среда влияет на формирование индивидуальности человека, сколько сама индивидуальность (с ее уникальной генетической конституцией) формирует свою особую среду. Впервые на это обстоятельство обратила внимание американская исследовательница Сандра Скарр в 1983 г.
Наличие генотип-средовой ковариации приводит к тому, что средовые условия у более близких родственников, имеющих высокую долю общих генов (т.е. сходные генотипы), оказываются более сходными, чем у более отдаленных родственников. Характерным примером могут служить идентичные близнецы, разлученные в раннем детстве и никогда не встречавшиеся друг с другом. Уже будучи солидными людьми, при первой встрече они с удивлением обнаруживают множество совпадающих обстоятельств своей жизни: сходство профессий, привычек, любимых занятий и даже сходные особенности спутников жизни.
Можно ли оценить вклад генотип-средовой ковариации в изменчивость признака? В психогенетике для этого существуют специальные методы. Большинство из них используют приемы, основанные на сравнении оценок характеристик среды у родственников различной степени родства. Одним из наиболее распространенных является методика интервью и наблюдений НОМЕ (Home Observation of Measurement of the Environment), позволяющая оценить эмоциональные и вербальные реакции родителей, отношение к наказаниям, организацию физической среды в семье и т.п. Используются также всевозможные опросники, в которых родственники оценивают различные параметры семейной и внесемейной среды. Полученные оценки могут сравниваться у родственников подобно тому, как это делается для любых количественных характеристик фенотипа. Так можно оценить сходство родственников не только по фенотипу, но и по средовым параметрам. Если средовое сходство оказывается выше у более близких родственников, можно предполагать наличие генотип-средовой ковариации.
Вопросы для закрепления:
Что такое ассортативность? Приведите примеры.
Приведите примеры дискретной и континуальной изменчивости.
Какой вид изменчивости характерен для большинства психологических признаков?
Как графически изображается частота встречаемости в популяциях для качественных и количественных признаков?
Что такое признаки с пороговым эффектом?
Приведите примеры качественных, количественных признаков и признаков с пороговым эффектом.
Как будет выглядеть частотное распределение для признака с пороговым эффектом?
В чем специфика понятия популяции в генетике?
Назовите основные критерии для отнесения сообществ организмов к популяции.
Каковы причины образования популяций?
Почему отдельные признаки встречаются в популяциях с разной частотой?
Какие изменения могут происходить в популяциях?
В чем специфика человеческих популяций?
Какие виды человеческих популяций вы знаете?
Какие признаки относятся к континуальным, а какие к дискретным?
Индекс массы тела - «ожирение».
Артериальное давление – «гипертония».
Плотность скелета – «перелом».
Сахар крови – «диабет».
Реактивность бронхов – «астма».
Нейротицизм – «тревога/депрессия».
Способность к чтению – «дислексия».
От каких факторов зависит количественная изменчивость признаков?
Объясните разницу между понятиями генотип, геном и генофонд.
Как можно объяснить, что генотипические и средовые факторы влияют на количественную изменчивость в популяции?
Как можно представить взаимодействие генотипа и среды в индивидуальном развитии человека?
Почему в психогенетике следует различать взаимодействие генотипических и средовых факторов при формировании популяционного разнообразия и при формировании индивидуального фенотипа?
Какие генотипы будут представлены в популяции, если ген имеет три аллельные формы?
Каковы будут фенотипы, если эти аллели определяют разную количественную выраженность признака, а эффекты доминирования отсутствуют?
Равномерно ли распределены генотипы в популяции? Покажите это на модели.
Что доказывает разную чувствительность генотипов к среде? Продемонстрируйте на модели.
От чего зависит количественная изменчивость в пределах одного генотипа?
От чего зависит фенотипическая изменчивость во всей популяции?
Почему дисперсия в пределах одного генотипа является чисто средовой?
Из чего слагается генетическая дисперсия в популяции?
Какие дисперсии в приведенной модели могут быть вычислены непосредственно?
Каким образом может быть вычислена генетическая составляющая дисперсии?
Какие составляющие популяционной фенотипической дисперсии выделяют?
Литература: [1, 6, 9, 10, 11, 12, 13,17,18].