

Циклическое и нециклическое фотосинтетическое фосфорилирование
Фотосинтетическое фосфорилирование - образование АТФ в ходе реакций, активируемых светом в хлоропластах, может осуществляться циклическим и нециклическим путями.
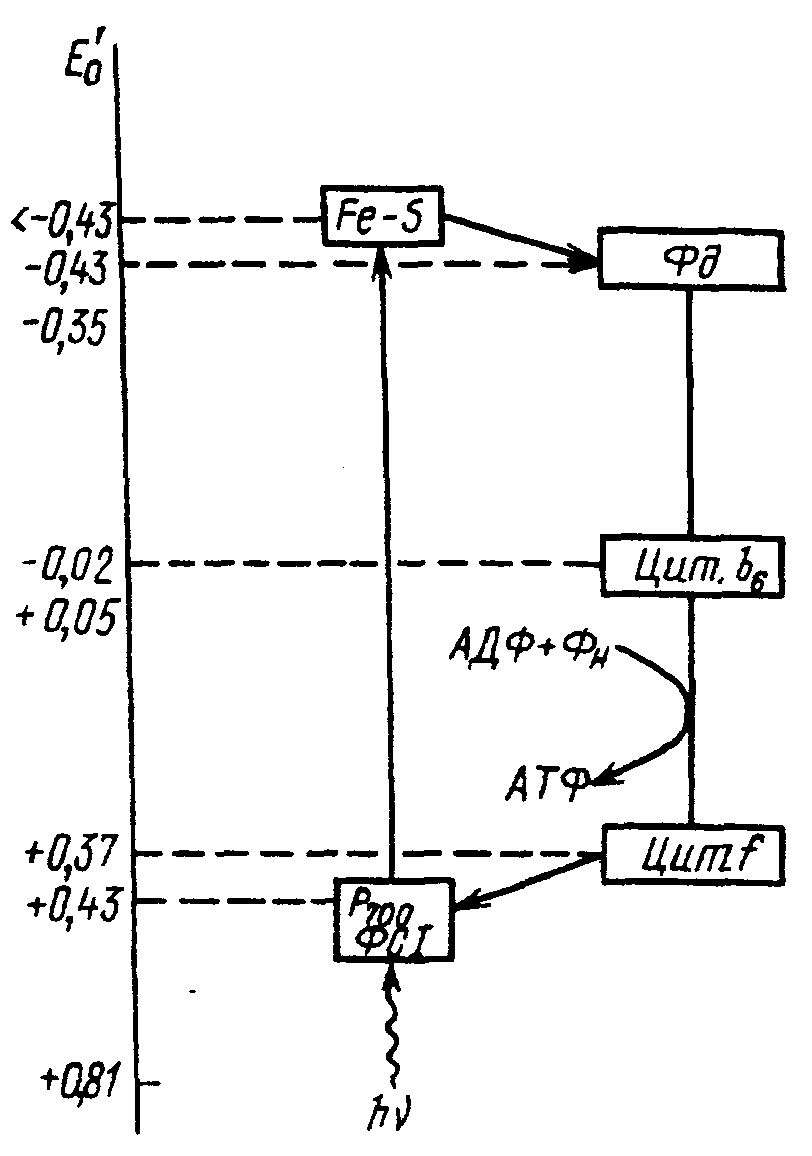
Рис. Схема циклического транспорта электронов.
При поглощении кванта света один их электронов пигмента РЦ (P700) переходит на более высокий энергетический уровень. В этом состоянии он захватывается белком, содержащим Fe и S (Fe-S-центр), а затем передается на Fe-содержащий белок ферридоксин. Дальнейший путь электрона - поэтапный, его транспорт обратно к P700 через ряд промежуточных переносчиков, среди которых имеются флавопротеиды и цитохромы. По мере транспорта электрона его энергия высвобождается и используется на присоединение Фн к АДФ с образованием АТФ.
Т.о., при циклическом фотофосфорилировании энергия света расходуется на перенос электрона на высоковосстановленное соединение, т. е. против градиента ОВ-потенциала, а затем электрон «скатывается с горки», образованной переносчиками с убывающим уровнем восстановленности, и, обедненный энергией, возвращается на ФС1. Механизмы сопряжения (АТФ-азный комплекс) обеспечивают на этом отрезке пути запасание энергии в виде АТФ. У простейших автотрофов - бактерий - это единственный путь фотофосфорилирования.
Нециклическое фотофосфорилирование (НЦФФ). У высших растений в процессе эволюции появился более сложный путь, который осуществляется при участии двух ФС и обеспечивает восстановление НАДФ за счет фотоокисления воды. Причем восстановление НАДФ осуществляет ФС 1, а фотоокисление воды - ФС 2. Эти две системы функционируют одновременно и взаимосвязанно. Р. Хиллом и Ф. Бендаллом (1960) разработана схема последовательности реакций, которая получила название схемы нециклического транспорта электронов, или Z-схемы (рис.).
При возбуждении P700 в РЦ ФС1 электрон захватывается мономерной формой хлорофилла а и затем последовательно передается через железосерные белки, ферридоксин, флавопротеиды на восстановление НАДФ. P700, не получив электрона обратно, как в случае ЦФФ, приобретает положительный заряд, который компенсируется электроном ФС2.
В ФС2 P680, возбужденный квантом света, передает электрон феофитину. От феофитина электрон, теряя энергию, последовательно передается на пластохиноны, железосерный белок, цитохром f, пластоцианин и, наконец, на P700 ФС1. Энергия, освобождающаяся при транспорте электрона от возбужденной ФС2 на ФС1, используется для синтеза АТФ из АДФ и Фн.
P680, оставшись без электрона, приобретает способность получать электрон от воды. Несмотря на активное исследование, детально механизм процесса фотоокисления воды не установлен. Показано участие белкового комплекса и переносчика электронов Z для функционирования которых необходимы Mn, Cl и Са.
Т.о., при НЦФФ происходит линейный или открытый (не замкнутый по циклу) транспорт электронов. Донором электронов является вода, конечным акцептором - НАДФ. Причем происходит одновременно двухэлектронный транспорт. Передача электронов осуществляется при участии двух ФС, поэтому для переноса каждого электрона расходуются два кванта света. На участке между ФС2 и ФС1 транспорт электрона идет по убывающему градиенту ОВП с высвобождением энергии и запасанием ее в АТФ.
Наряду с нециклическим в мембранах хлоропластов высших растений функционирует циклический транспорта электронов. Причем ферридоксин выполняет роль регулятора потока электронов. При возрастании потребности в АТФ часть электронов от ферридоксина через систему цитохромов возвращается к P700 с образованием АТФ. Восстановление НАДФ в этом случае не идет, и фотоокисления воды при участии ФС2 нетребуется.
Белковые комплексы, участвующие в фотохимических реакциях фотосинтеза, в мембранах хлоропластов распределены неравномерно. Наибольшие различия наблюдаются между теми мембранами, которые плотно состыкованы друг с другом в гранах и теми участками мембран, которые контактируют со стромой, - закругленные участки тилакоидов гран и мембраны тилакоидов стремы (рис.).
ФС2 и ССК находятся в основном в плотносостыкованных мембранах гран. ФС1 со своим ССК и АТФ-азный комплекс находятся на несостыкованных участках мембран. Взаимодействие между ними осуществляется с помощью легкоподвижного в липидной фазе пластохинона, а также благодаря перемещению водорастворимого пластоцианина вдоль внутренних поверхностей ламелл и водорастворимого ферридоксина вдаль наружных поверхностей.
К латеральным перемещениям способны пигментбелковые комплексы. Например, восстановление в избыточных количествах пластохинона ФС2 способствует перемещению ССК в стромальную область мембраны и возрастанию миграции поглощенной энергии света с ФС1. Возросшая фотохимическая активность ФС1 усиливает окисление пластохинона, что приводит к обратному перемещению ССК. Таким образом, система взаимной регуляции активности ФС1 и ФС2 включает латеральный транспорт белковых комплексов в мембранах хлоропластов.