

Почки, метабоизм веществ в почках
..docНеобходимое условие жизнедеятельности - постоянство внутренней среды. К ее параметрам относятся, в частности, общее содержание воды в организме и соотношение объемов водных пространств, зависящее от гидростатического и осмотического давления. Для поддержания постоянства этих параметров необходимо точное соответствие выведения воды и осмотически активных веществ их потреблению. Эта задача усложняется тем, что прием твердой и жидкой пищи постоянно колеблется. Хотя выделительная функция отчасти осуществляется ЖКТ, легкими и кожей , главные органы выведения - это почки. В процессе клеточного метаболизма в почках образуется моча, которая является основным экскретом организма.
Почки регулируют:
-
водно-электролитный обмен,
-
кислотно-основное равновесие,
-
выведение из организма конечных продуктов метаболизма,
-
осмотическое давление,
-
стимулируют эритропоэз.
Почки наделены уникальными экскреторными и внутрисекреторными функциями, сопряженными с расходыванием энергии АТФ. Почки млекопитающих могут использовать в качестве субстратов для получения АТФ жирные (пальмитиновая и т. п.) и другие карбоновые кислоты (молочная и т. п.), моносахариды (глюкозу и т. п.), кетоновые тела. Основным источником для получения АТФ в почках является аэробный путь окисления субстратов.
Экскреторная функция почек.
Основной расход АТФ в почках связан с процессом реабсорбции NaCI почечными канальцами во внеклеточные пространства. Кора и мозговой слой почек имеют значительные различия в обмене веществ. Кора почек имеет высокую степень потребления кислорода для цикла Кребса и реакций окислительного фосфорилирования. Основным источником энергии для коры почек являются жирные кислоты. Окисление жирных кислот при этом сопровождается образованием кетоновых тел. Мозговой слой почек имеет высокую концентрацию ферментов анаэробного гликолиза, что связано с низким уровнем реакций цикла Кребса, небольшим потреблением кислорода и энергии. Основным источником энергии в мозговом слое почек является глюкоза плазмы крови после приема корма животным.
За счет глюконеогенеза под контролем адреналина и глюкагона кора почек способна синтезировать глюкозу быстрее, чем печень, но почки не поставляют значительных количеств глюкозы в кровяное русло. При этом глюкоза, синтезируемая в коре почек, служит важным источником энергии и для мозгового слоя.
В почках имеется активный фермент Na+-К+-АТФаза, который гидролизует АТФ в присутствии натрия и калия с использованием высвобождающейся энергии на активный пере нос натрия в клетках:
3 Na+
+ 2К+ + АТФ + Н2О 3Na+ + 2К+ + АДФ +
Фн
Na+
+ 2К+ + АТФ + Н2О 3Na+ + 2К+ + АДФ +
Фн
Экскреторная функция почек хорошо согласуется с интенсивным их кровоснабжением. Отметим наличие в почках клубочкового и канальцевого аппарата, весьма различных по своей функции и в то же время тесно связанных между собой. Кровь, находящаяся в сосудистых клубочках, отделена от содержимого канальцев трехслойной мембраной, которая образует своеобразный фильтр спорами ультрамикроскопических размеров. Через этот фильтр могут проходить вода, все низкомолекулярные компоненты плазмы крови и даже некоторые пептиды плазмы крови в небольшом количестве. Это позволяет считать, что первичная моча, поступающая в каналец, представляет собой ультрафильтрат крови (т. е. все главные компоненты крови, но без белков). Концентрация натрия, калия, фосфатов, глюкозы, мочевой кислоты, креатинина в первичной моче практически такая же, как и в плазме крови. При патологических состояниях фильтр может пропускать в каналец и некоторые белки. Наличие белка в моче может быть связано не только с повреждением клубочков, но и с нарушением структуры и функции канальцев.
Состав клубочкового фильтрата, как было отмечено, определяется составом безбелковой части плазмы крови. Количество его зависит от величины кровяного давления в почечных капиллярах, которому противодействует онкотическое давление белков плазмы и внутрикanсулярное давление.
Основное количество ультрафильтрата подвергается обратному всасыванию в канальцах. Выполняемая ими работа по обратному всасыванию очень велика, что, естественно, требует очень хорошего энергетического снабжения. Интенсивность окислительных процессов в почках подтверждается высокой потребностью органа в кислороде. Почки поглощают до 10% всего потребляемого организмом кислорода.
Почечный каналец подразделяется на проксимальный и дистальный сегменты. В проксимальном сегменте происходит реабсорбция примерно 90% воды и 80% ионов натрия и хлора. Здесь же почти полностью реабсорбируется глюкоза, аминокислоты и небольшие количества пептидов, содержащиеся в ультрафильтрате, а также фосфаты и бикарбонаты. Всасывание воды в проксимальном сегменте происходит пассивно. Всасывание натрия идет активно против градиента концентраций (в содержимом канальца концентрация натрия ниже концентрации его в крови). В клетках канальцев происходит и всасывание большей части калия. Мочевина, мочевая кислота плохо всасываются в канальцах и поэтому значительная часть их из фильтрата переходит во вторичную мочу.
Рассмотрим теперь роль почек в поддержании постоянства кислотно-основного равновесия организма. Выше было показано, что в организме образуется много кислых продуктов метаболизма. Выведение их только в форме катионов (Na+, К+ и др.) привело бы к значительному снижению содержания последних в организме. Однако почки располагают несколькими механизмами, позволяющими, с одной стороны, обеспечить выведение кислот, а с другой - сохранить необходимые для организма катионы.
Одним из таких механизмов является возможность выведения слабых кислот как таковых. Содержащиеся в фильтрате анионы кислот соединяются с водородом, образующимся в процессе диссоциации угольной кислоты (Н2СОз ~ н+ + НСОз -) с образованием слабодиссоциирующих карбоновых кислот.
Источником угольной кислоты является реакция взаимодействия воды и углекислого газа при участии фермента карбоангидразы, которым клетки почечных канальцев весьма богаты. Образующийся наряду с протоном бикарбонатный ион всасывается обратно в кровь. Характерно, что изучение этого процесса позволило использовать ингибиторы карбоангидраз в качестве мочегонных препаратов.
Однако сильные кислоты выделяться с мочой как таковые не могут. В этом случае для экономии натрия используется другой, менее важный катион - ион аммония (NH4 +). Источником этого иона является процесс дезаминирования аминокислот, и прежде всего - глутамина. В результате такого дезаминирования образуется аммиак которыи В дальнеишем соединяется с протоном, освобождающимся при диссоциации угольной кислоты:
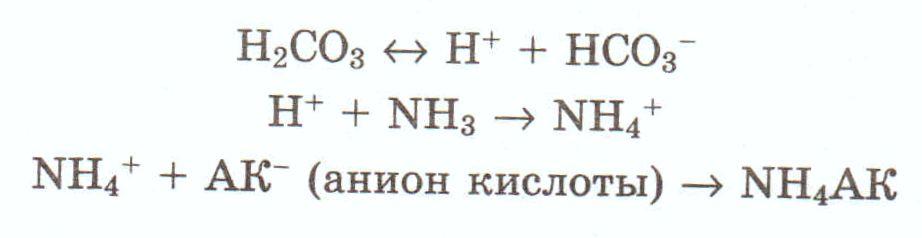
Таким образом, участвуя в удалении из организма кислых продуктов обмена веществ, почки одновременно выполняют и сберегающую натрий функцию. Экономия натрия достигается и за счет превращения Na2HP04 в NaH2P04•
В плазме крови, и, следовательно, в первичной моче, преобладает двухосновной фосфат (Na2HP04). Под влиянием иона водорода, секретируемого клетками почечных канальцев в просвет, образуется одноосновной фосфат _ NaH2P04, тогда как один ион натрия из просвета поступает обратно в кровь. Следовательно, в основе серии этих процессов лежит секреция ионов водорода и реабсорбция ионов натрия и бикарбоната.
Внутрисекреторная функция почек связана с синтезом белка ренина, обладающего протеолитическим действием. Субстратом в реакции с участием ренина является белок ангиотензиноген. Из ангиотензиногена плазмы крови образуется ангиотензин 1, состояrций из 10 аминокислот. С участием специальной пептидазы сыворотки крови ангиотензин 1 превраrцается в ангиотензин 11, содержаrций в молекуле на две аминокислоты меньше. Ангиотензин 11 оказывает влияние на надпочечники, стимулируя выделение альдостерона. Последний повышает реабсорбцию натрия и воды в канальцах, что приводит к изменению кровяного давления. Полагают, что повышенное образование ренина является важной причиной развития гипертонии.
Почки также стимулируют эритропоэз. Пролиферативная активность эритропоэза контролируется эритропоэтином почек. Синтезируемый в печени профактор (ф-глобулин) транспортируется в почки, где он превращается в активный эритропоэтин.
При должном уровне эритропоэтина в почках синтез гемоглобина в костном мозге протекает активно.
Относительно стабильное осмотическое давление крови поддерживается за счет баланса между поступлением воды с питьем и пищей и потерей воды с выдыхаемым воздухом, потом, калом и мочой. За тонкую регуляцию осмотического давления отвечает антидиуретический гормон , путем изменения проницаемости дистальных извитых канальцев в почках и собирательных трубок в почках .
При недостаточном потреблении воды , сильном потоотделении или после приема большого количества соли осморецепторы, находящиеся в гипоталамусе , регистрируют повышение осмотического давления крови . Возникают нервные импульсы, которые передаются в заднюю долю гипофиза и вызывают высвобождение антидиуретического гормона, который повышает проницаемость для воды стенок дистального извитого канальца и собирательной трубки, вода выходит из фильтрата в тканевую жидкость коркового вещества почек и мозгового вещества почек , и почки выделяют меньший объем более концентрированной мочи .
Антидиуретический гормон повышает также проницаемость собирательной трубки для мочевины , которая диффундирует из мочи в тканевую жидкость мозгового вещества почки, в результате чего повышается осмолярность и происходит увеличение выхода воды из тонкого сегмента восходящего колена петли Генле .
После приема большого количесва воды осмотическое давление крови уменьшается и секреция антидиуретического гормона прекращается. Стенки дистального извитого канальца и собирательной трубки становятся непроницаемыми для воды, реабсорбция воды при прохождении фильтрата через мозговое вещество почки уменьшается и, следовательно, выводится большой объем гипотонической мочи.
При недостаточности антидиуретического гормона возникает несахарный диабет , при котором выделяется очень большое количество гипотонической мочи.
Моча.
В моче животных содержится свыше 200 различных компонентов, относящихся как к органическим, так и к минеральным веществам. Количество суточной мочи у разных видов животных колеблется в широких пределах и часто зависит от, характера кормления, количества выпитой жидкости, физической нагрузки на организм. Количество мочи в сутки составляет у коров 6-20 л, у овец - 1-2 л, у лошадей - 3-10 л, у свиней - 2-6 л, у собак - 0,5-2 л, у кошек - 0,2-0,3 л.
Плотность мочи колеблется в пределах 1,02-1,06. У травоядных животных моча имеет слегка щелочную реакцию, а у плотоядных - кислую. При смешанном рационе реакция мочи изменяется в зависимости от преобладания растительных кормов или кормов животного происхождения. Моча образуется из компонентов плазмы крови, однако отличия их по составу весьма значительны. В частности, моча не содержит в норме таких компонентов плазмы крови, как белки, полипептиды, глюкоза.
Экскреторная функция почек находится под контролем центральной нервной системы, гормонов гипофиза и надпочечников. Поэтому болевые ощущения и стрессовые ситуации могут рефлекторно вызвать анурию (прекращение образования мочи), тогда как болезни ЦНС, гипофиза и надпочечников могут привести к полиурии (повышенное выделение мочи).
Органические компоненты мочи представлены в основном азотсодержащими веществами - продуктами обмена белков. В моче содержатся мочевина, мочевая кислота, аммиак, пуриновые основания, креатинин; в небольшом количестве - производные продуктов гниения белков в кишечнике (индол, скатол фенол).
Среди органических соединений небелкового происхождения в моче встречаются соли щавелевой кислоты, молочной кис лоты, кетоновые тела. Глюкозы в моче в обычных условиях не должно быть.
Эритроциты появляются в моче (гемотурия) при заболеваниях почек и мочевыводящих органов. В моче содержатся пигменты (уробилин, урохром), которые и определяют цвет мочи. С мочой выделяются электролиты (Na+, К+, С1-, Са2+, Мg2+, сульфаты и др.). В моче содержатся гормоны и их метаболиты, ферменты, витамины.
Основным азотсодержащим веществом мочи млекопитающих является мочевина. На ее долю приходится более 80% от общего азота мочи. При скармливании рационов, богатых белками, количество выделяемой мочевины значительно увеличивается. Лишь у птиц основным продуктом обмена белков является мочевая кислота. В моче жвачных животных содержится в небольших количествах и мочевая кислота, образующаяся при распаде пуриновых оснований. у птиц распад пуриновых оснований на этом компоненте и заканчивается, но все другие виды животных способны далее расщеплять мочевую кислоту и выделять продукт ее окисления - аллантоин. У рыб выделение продуктов распада пуриновых оснований происходит в форме аллантоиновой кислоты.
В моче животных содержится креатин и креатинин, который является ангидридом креатина:
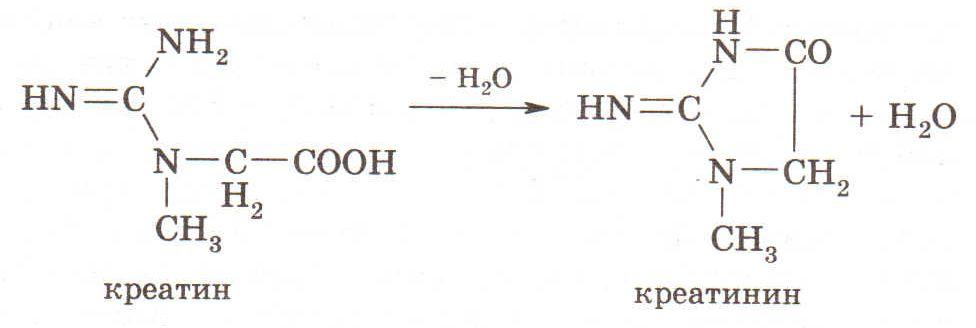
Креатин играет основную роль в метаболизме макроэргических фосфатов. Креатин (N-метилгуанидиноацетат) является производным глицина, аргинина и S-аденозилметионина.
Креатинин образуется путем отщепления воды из креатина или путем отщеплен ия фосфорной кислоты от фосфорилированной формы креатина - креатинфосфата. Количество креатинина в моче зависит от функциональной активности мышечной ткани, где креатинфосфат обеспечивает энергией мышечное сокращение. В норме количество выделяемого с мочой креатинина значительно больше, чем креатина, и лишь при распаде мышечной ткани выделение самого креатина увеличивается. Это отмечают при мышечной дистрофии, вызванной недостаточностью витамина Е.
Креатинин фильтруется через базальную мембрану клубочков и в норме не реабсорбируется в тубулярном отделе нефрона. В условиях повышенной концентрации креатинина в крови часть его активно экскретируют клетки тубулярного эпителия. В норме образование креатинина и его выведение эквивалентны. Повышение уровня креатинина и мочевины в крови – признак почечной недостаточности. Однако повышение концентрации креатинина при почечной недостаточности происходит раньше, чем повышение концентрации мочевины. Определение содержания креатинина в крови и моче используют для оценки скорости клубочковой фильтрации. В клинической практике наиболее распространенной является проба Реберга. В действительности точное измерение скорости клубочковой фильтрации требуется редко. Показаниями для ее определения могут быть: обследование пациентов с минимальными нарушениями почечной функции, обследование потенциального донора почки, определение исходной дозировки потенциально токсичного лекарства, выводимого через почки. Для большинства больных с установленной болезнью почек для оценки их функции достаточно серийных измерений креатинина в плазме.
Проба Реберга основана на определении креатинина в крови и моче (клиренс креатинина - отношение уровней креатинина в моче и плазме, умноженное на минутный диурез). До начала выполнения теста необходимо прекратить лечение кортикотропином, кортизолом, тироксином, избегать физических нагрузок. Обследуемый натощак выпивает 400-500 мл воды и мочится. Эту порцию мочи выливают, время мочеиспускания точно отмечают. Через полчаса пунктируют локтевую вену пациента и получают кровь. Еще через полчаса (через час после мочеиспускания) собирают мочу. По объему собранной мочи устанавливают минутный диурез, в крови и моче определяют концентрацию креатинина, по формулам рассчитывают фильтрацию и реабсорбцию. Клиренс креатинина у взрослых в норме составляет 80-120 мл/мин, реабсорбция -97-99%. Для корректного расчета клиренса креатинина важно знать рост, вес и возраст пациента.
Моча содержит и небольшое количество стеркобилина. Стеркобилин образуется в толстом кишечнике, откуда попадает в систему кровообращения и далее в мочу. С мочой выделяются и различные аминокислоты, количество которых увеличивается при нарушении функции печени в связи с уменьшением способности этого органа к их дезаминированию.
Моча содержит также парные соединения, образующиеся в печени при обезвреживании некоторых токсических для организма веществ. К их числу относятся гиппуровая кислота, индикан. Гиппуровая кислота представляет собой соединение бензойной кислоты с гликоколом.
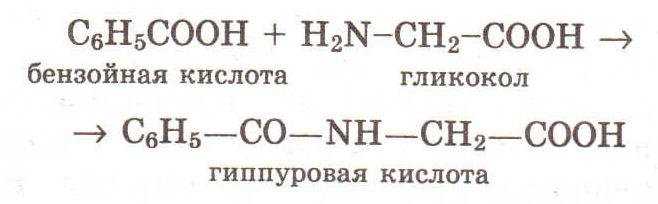
Гиппуровая кислота в значительных количествах находится в моче травоядных животных. Так, в моче лошади в сутки выделяется около 160 г этой кислоты. У птиц бензойная кислота соединяется не с гликоколом, а с аминокислотой орнитином, образуя орнитуровую кислоту.
Из безазотистых компонентов в моче постоянно присутствуют ряд органических кислот, таких как щавелевая, молочная, лимонная, масляная, ацетоуксусная, В-гидроксимасляная. Моча содержит небольшое количество индоксилсерной кислоты (индикан). Она образуется из индола, освобождающегося из триптофана в толстом кишечнике животного под влиянием гнилостной микрофлоры. В норме ее количество в моче невелико, а увеличение наблюдается при нарушении моторики кишечника. Количество индикана в моче свидетельствует о скорости гниения белков в кишечнике и о функциональном состоянии печени.
На долю минеральных соединений мочи приходится около 40% ее плотного остатка. Анионы мочи представлены хлором, остатками фосфорной и серной кислот. Количество хлоридов мочи зависит от их потребления и подвержено в связи с этим значительным колебаниям. Снижение количества хлоридов в моче наблюдается при рвотах, вследствие потери соляной кислоты желудочного содержимого. Фосфор мочи содержится в форме солей ортофосфорной кислоты, в основном NaН2P04 И Na2HP04' Сера выделяется в форме сульфатного иона, а также в эфирной форме, например, в форме соединения фенола с серной кислотой. Катионы главным образом представлены натрием, калием, кальцием, магнием и аммонийным ионом. Выделение с мочой натрия и калия превышает выделение кальция и магния, так как щелочноземельные элементы в значительной степени выделяются из организма желудочнокишечным трактом. Из неорганических соединений в моче в наибольшем количестве содержится хлористый натрий.
Количество аммонийного иона в моче (NH+) в значительной степени зависит от состояния кислотно-основного равновесия. Выше уже было изложено значение этого иона для связывания анионов кислот. Значительное увеличение концентрации аммонийных ионов (аммиака мочи) наблюдается при ацидозах.
Признаком патологии может быть обнаружение компонентов, которых в норме в моче животных нет, или же увеличенное содержание обычно содержащихся веществ.
В моче здоровых животных белок в норме отсутствует. То небольшое количество полипептидов, которое может выделяться, не обнаруживается обычно используемыми в лабораторной практике качественными реакциями. Наиболее простым способом обнаружения являются реакции, основанные на осаждении, белка концентрированными кислотами (сульфосалициловая, азотная кислоты). Обнаружение белка, главным образом альбуминов, в моче всегда указывает на поражение почек или мочевыводящих путей. Это может быть белок плазмы крови, попадающий в первичную, а затем и во вторичную мочу из плазмы крови при повышении проницаемости мембран клубочков почек. Кроме того, белок в моче встречается и при поражении канальцевого аппарата. Часто наличие белка в моче (протеинурия) сопровождается и выделением форменных элементов крови, в первую очередь эритроцитов.
Ничтожное количество глюкозы, выделяемое с мочой здоровым организмом (10-20 мг на 100 мл), не обнаруживается используемыми в клинической практике методами. Выделение глюкозы с мочой (глюкозурия) может быть алиментарного (кормового) характера, однако это встречается редко. Разовая нагрузка глюкозой при водит к значительному повышению содержания глюкозы в моче. В связи с этим в первичную мочу фильтруется большее, чем в норме, количество глюкозы и канальцы не успевают обеспечить обратное всасывание сахара в кровь. В результате не всосавшаяся обратно глюкоза попадает во вторичную мочу. Более частой причиной глюкозурии является сахарный диабет, вызванный недостаточной концентрацией инсулина в крови.
В отсутствие инсулина глюкоза плохо проходит через клеточные мембраны, концентрация ее в крови резко возрастает и далее наблюдается та же картина, что и при алиментарной глюкозурии. Наличие глюкозы в моче встречается и при заболеваниях почек, когда фильтруется из крови нормальное количество глюкозы, но пораженные патологическим процессом почечные канальцы теряют способность к обратному всасыванию глюкозы из первичной мочи в кровь. Отличить почечную глюкозурию от глюкозурии при сахарном диабете можно путем определения концентрации глюкозы в крови. При диабете концентрация глюкозы в крови повышена, а при почечных заболеваниях она остается нормальной.
Одним из легко определяемых при патологических состояниях компонентом мочи являются кетоновые тела (ацетон, ацетоуксусная и гидроксимасляная кислоты). Кетоновые тела - нормальный компонент крови, и, следовательно, в числе других компонентов плазмы крови они содержатся в первичной моче в незначительном количестве, которое не обнаруживается качественными реакциями, используемыми при лабораторном исследовании мочи. Повышенное количество кетоновых тел в моче (кетонурия) наблюдается при кетозах животных и при сахарном диабете. Богатые жиром и белком концентраты дают при своем распаде в· организме большое количество молекул ацетил-КоА, не успевающего распадаться в цикле Кребса. Тем самым создаются благоприятные условия для синтеза из ацетилкоэнзима А кетоновых тел. Повышенное количество кетоновых тел в моче имеет место и в условиях недостаточного кормления животного, когда организм получает энергию за счет распада собственных жиров организма. При диабете вследствие нарушения проникновения глюкозы из крови в клетки, последние испытывают энергетический голод и вынуждены получать энергию за счет почти исключительно распада жиров, а это, в свою очередь, приводит к накоплению в организме молекул ацетилкоэнзима А и превращению его в кетоновые тела
К патологическим компонентам. мочи относятся также желчные пигменты и уробилин. Через почечные мембраны может проходить только прямой билирубин, а он образуется только в печени. Патология паренхимы печени сопровождается распадом части печеночных клеток, повышением проницаем ост и клеточных мембран этого органа. В результате содержащийся в клетках прямой билирубин поступает в кровь и с кровью в почки, где путем фильтрации попадает в первичную, а затем во вторичную мочу. Уробилин является продуктом превращения билирубина, его восстановления. Превращение билирубина в уробилиноген происходит в тонком кишечнике. Большая часть уробилиногена выделяется с калом, 10-20% его реабсорбируется из кишечника по воротной вене в печень, где и разрушается. Около 80% реабсорбированного уробилиногена повторно выделяется печенью в кишечник. Однако патология печени не позволяет в полной мере осуществить этот процесс, и содержащийся в клетках уробилиноген вследствие повышения проницаемости мембран клеток печени поступает в кровь и далее в мочу.
Концентрация уробилиногена в моче, таким образом, зависит от многих факторов, в частности, от количества билирубина, поступающего с желчью в кишечник, от способности кишечной флоры превращать билирубин в уробилиноген, от количества всосавшегося в кишечнике уробилиногена, от способности печени поглощать его из крови после всасывания в кишечнике, от состояния функции почек.
Следует отметить, что моча содержит и в норме небольшое количество продуктов превращения желчных пигментов в кишечнике, и в частности, стеркобилиногена, сходного, в определенной степени, по своей химической структуре с уробилиногеном. Однако при заболеваниях печени количество этих продуктов в моче значительно увеличивается. При закупорке желчных протоков в моче обнаруживается лишь прямой билирубин, а уробилин отсутствует, так как желчь не поступает в кишечник. При гепатите моча содержит повышенное количество билирубина и уробилиногена. Повышенное содержание уробилиногена в моче отмечено при снижении моторики кишечника и при сдвиге рН мочи в щелочную сторону.
При длительном застое кормовых масс в толстом кишечнике, что сопровождается усилением гнилостных процессов и распадом циклических аминокислот, в кровь поступает значительное количество индола и скатола, а также фенола и крезола. Эти токсичные для организма вещества с кровью поступают в печень, где обезвреживаются, соединяясь с серной или глюкуроновой кислотой И превращаясь в соответствующие парные кислоты - индоксилсерную или индоксилглюкуроновую кислоты И т. д. Калиевая соль индоксилсерной кислоты (индикан) определяется в моче с помощью специфической качественной реакции. Выраженная реакция на индикан указывает на усиление в организме гнилостных процессов. К патологическим образованиям следует отнести и мочевые камни, выпадающие в почках вследствие нарушения сбалансированного кормления, понижения растворимости солей, связанной с изменением концентрации водородных ионов в моче, малой двигательной активностью. Обычно камни имеют сложный состав, однако их принято делить на образованные солями фосфорной кислоты, щавелевой или мочевой кислоты. В значительной степени они представлены кальциевыми солями. У птиц, у которых мочевая кислота является основным продуктом азотистого обмена, избыток белка в рационе приводит к образованию очень большого количества мочевой кислоты, которая будучи плохо растворимым соединением, может полностью закупорить мочевые пути. Отметим, что моча у птиц выделяется в клоаку, где смешивается с каловыми массами.
Литература.
-
http://www.labdiagnostic.ru/docs/specialists/bioxim_pokazat.shtml
-
С. Ю. Зайцев, Ю. В. Конопатов «Биохимия животных» СПб: Издательство «Лань» 2005
-
Т.Т. Березов, Б.Ф. Коровкин "Биологическая химия", Москва, "Медицина", 1998 год
-
А. С. Чиж «Нефрология в терапевтической практике» http://www.uroweb.ru/catalog/med_lib/nefrologij/book.php