

Лекции по биофизике
.pdfIX. БИОФИЗИКА СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
Общие положения
Различают несколько видов синаптической передачи (синапсов):
1.Электрические синапсы.
2.Химический синапс.
3.Смешанные синапсы.
Их функция обеспечить (возбуждающие синапсы) или предотвратить (тормозные синапсы) передачу нервного импульса с одного электровозбудимого объекта (нейрон–нейрон, нейрон–мышца) на другой.
Критерием дифференцировки синапсов являются их отличия касательно электрофизиологических свойств, морфологических характеристик, фармакологической регуляции.
А. Электрофизиологические свойства:
1. Наличие электрической связи между клетками Для электрического синапса (Э.С.) она обеспечивает зависимость тока
входящего направления через мембрану пресинаптической клетки от:
Э.С. |
Vпостсинапт. |
и равна 0,1-0,5 (определяется RC- - |
|
Vпресинапт. |
мембран); |
||
|
Для химического синапса (Х.С.) |
0. |
2. Величина синаптической |
задержки (интервал между началом |
пресинаптического ПД и постсинаптической деполяризацией): Для Х.С. он равен 0,2 и выше (до 0,8-100)мс.
Для Э.С. = 0,1 мс. (зависит от RC- -мембран).
3. Зависимость постсинаптического потенциала (ПСП) от исходного уровня мембранного потенциала (Vм):
Для Э.С. – нет зависимости, так как ПСП определяется Vпресинапт. и коэффициентом передачи.
Для Х.С. – есть и величина тока Ii через постсинаптическую мембрану равна:
Ii gi (Vi Vm ) ,
где: Vi и Vm – уровни ПСП и исходного мембранного потенциала
Б. Морфологические характеристики:
Э.С. отличает малая ширина синаптической щели (100 нм), наличие специализированных структур щелевых контактов. В их состав входят особые каналы, проницаемы для низкомолекулярных веществ (1-2 кД). Щелевые контакты получили название высокопроницаемых контактов
121

(ВПК). Они чувствительны к механическому разобщению из-за удаления ионов кальция из раствора, алкалоза, действия гипертонического раствора.
Рис. 25. Химический синапс
Химический синапс представляет особую структуру, в состав которой входят пре- и постсинаптическая мембраны (толщина мембран 4-5нм) и синаптическая щель (100-150мкм). Пример-двигательная пластинка нервномышечной синаптической передачи. Зона активного контакта концевой пластинки -1,5 мкм. Аксоплазма имеет синаптические пузырьки диаметром
50 нм.
В. Фармакологические критерии:
Э.С. – не чувствительны к замене Са2+ на Mg2+, Со2+.
Х.С. – чувствительны к действию медиаторов и их метаболизму, уровню ионов кальция.
Электрические синапсы
Каждый из видов синаптической передачи имеет преимущества и недостатки. Например, Э.С. отличает от Х.С. то, что они:
1.малозависимы от метаболизма (ферментов);
2.нет истощения функций;
3.проводят высокочастотные серии импульсов.
Электрические синапсы подразделяют на:
1.Невыпрямляющие синапсы. Когда ток через постсинаптическую
мембрану вызывает линейные сдвигиVмембр. независимо от направления движения импульса.
2.Выпрямляющие. Когда Vpre Vpost и импульс пользуется преимуществом проведения в одном из направлений.
Химический синапс
Условиями функционирования Х.С.является следующее:
1.медиатор выделяется при возбуждении;
2.должен прийти стимул – эффект возбуждения;
3.блокаторы синаптической передачи должны быть активны;
4.должно быть метаболическое обеспечение медиатора.
Электрические явления в Х.С.:
1. В состояние покоя регистрируется миниатюрный потенциал концевой пластинки (МПКП) – электрический эквивалент выделения в синаптическую
122

щель спонтанного выделения 1 кванта медиатора. МКПК = 0,4-0,6мВ. Зона действия несколько каналов.
2. При стимуляция нейрона 30-50 Гц регистрируется ответ -50мв - потенциал концевой пластинки (ПКП). Он соответствует критическому уровню деполяризации скелетного мышечного волокна.
Квантово-везикулярная теория(Katz, Melodi).
Пусть m – квантовый состав ПКП; n – величина одного кванта; р – вероятность выделения кванта (m = pn); где m – целое случайное число и различие между ответами кратно n.
Для параметров n  400 и р
400 и р  0,25 эта теория верна. В ответ на изменения пресинаптического потенциала возникает кривая Ленгмюра зависимости от ПКП (
0,25 эта теория верна. В ответ на изменения пресинаптического потенциала возникает кривая Ленгмюра зависимости от ПКП (
Рис. 26).
Рис. 26. Зависимость
пресинаптического потенциала от постсинаптического
Как уже ранее утверждалось, основным электрофизиологическим отличием Х.С. от Э.С. является то, что амплитуда ПКП зависит от Vm скелетной мышцы. Причины такой зависимости в следующем.
1.Исходно амплитуда ПКП всегда на 10-20 мВ ниже ПД мышц. Причиной является постоянный шунт сопротивления мембраны КП, развивающийся под действием на нее нейромедиатора ацетилхолина (АХ). В результате высвобождения АХ в синаптическую щель снижается сопротивление постсинаптической мембраны в 1000 раз.
2.Мембраны КП и мышечного волокна по сути одна электрофизиологическая структура (Рис. 27).
123

Рис. 27. Эквивалентная
электрическая схема синапса
Если ключ К разомкнут, то регистрируется ПП мышцы (-80мВ). Замыкание К увеличивает проницаемость постсинаптической мембраны. Если е Е, то замыкание ключа вызывает ток в цепи е Е (е -15-0мВ). Ток бежит по первому пути и начинается деполяризация постсинаптической мембраны до уровня:
g
V g G е Е ,
где g и G – проницаемости мембран КП и мышечного волокна, соответственно. Естественно, чем ниже Е, тем выше V (отличие химического синапса).
В итоге:
1.При е = Е, ПКП = 0, уровень потенциала реверсии;
2.Если е 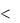 Е, ПКП меняет свой знак и наступает гиперполяризация постсинаптической мембраны – торможение передачи возбуждения на мышцу;
Е, ПКП меняет свой знак и наступает гиперполяризация постсинаптической мембраны – торможение передачи возбуждения на мышцу;
3.Так как присутствие См – шунтирует V, и Vm не будет совпадать с развитием постсинаптического мембранного потенциала (ПКП), спад ПКП относительно спада (тока ТКП) концевой пластинки запаздывает и ТКП является характеристикой действия медиатора.
4.Е = е при потенциале реверсии ( реверсии при Ер -15мВ). При этом уровне ПКП тока после действия медиатора не будет.
-15мВ). При этом уровне ПКП тока после действия медиатора не будет.
5.Величина потенциала реверсии рассчитывается по формуле:
|
ЕрWa |
Ер К |
|
реверсии |
2 |
, |
|
|
|||
где: ЕрNa и ЕрК – равновесные потенциалы для ионов натрия и калия, соответственно.
6. Нейромедиатор (АХ) одновременно увеличивает проницаемость для
Na+ и К+:
124
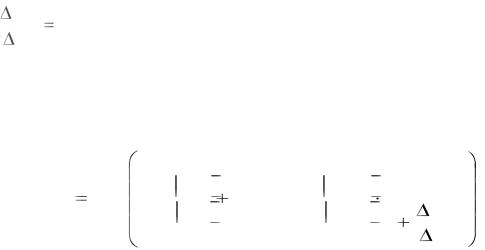
gNa |
1,29 =const на всем протяжении мембраны в сфере действия |
|
gK |
||
|
нейромедиатора.
Уравнение Takeuchi. Если величина потенциала постсинаптической мембраны зависит только от количества медиатора, то потенциал реверсии будет равен:
Е |
|
RT |
ln |
K ext |
|
Δg Na |
lg |
Na ext |
|
|
1 |
|
р |
F |
K int |
|
Δg K |
Na int |
|
|
g Na |
|
|||
|
|
|
|
|
|
|
||||||
|
|
|
|
|
1 |
|
||||||
|
|
|
|
|
|
|
|
|
g K |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
где: обозначения соответствующих величин концентраций, проводимости и констант объяснялись выше.
Основные положения о судьбе медиатора в химическом синапсе (Шеррингтон, 1897 г.)
1.В холинэргическом синапсе производится и утилизируется при действии одного импульса 2-200·103 молекул ацетилхолина (АХ). Возникает вопрос: «Откуда они берутся?».
2.Тем боле, что амплитуда МПКП определяется: размерами кванта медиатора, объѐмом диффузионного пространства, плотностью рецепторов в постсинаптической мембране, активность фермента расщепляющего АХ – ацетилхолинэстеразой (АХЭ), входным сопротивлением мембраны КП.
3.Оказывается, до взаимодействия с КП медиатор претерпевает достаточно сложные изменения:
a)Синтез и загрузку в везикулы (перикарион, тело нейрона);
b)Аксональный транспорт;
c)Слияние везикулы с пресинаптической мембраной и высвобождение медиатора в синаптическую щель;
d)Диффузия через синаптическую щель;
e)Узнавание, взаимодействие с рецепторами постсинаптической мембраны, изменение проницаемости ионов КП;
f)Расщепление АХЭ АХ.
g)Ресинтез ацетилхолина связан с активностью АцетилКоА, гликолизом холина из фосфотидилхолина:
АцетилКоА + холин – АХ (тело нейрона).
h) Загрузка синаптических пузырьков – это активный транспорт.
Концентрация АХ в них больше 5мМ.
j) Диффузия АХ от тела нейрона по аксону к месту высвобождения происходит с помощью олигосахаридного матрикса.
125
k) Высвобождение нейромедиатора связано с ионами кальция. Считают, что везикула его высвобождает прямо в синаптическую щель.
126
X. БИОФИЗИКА СОКРАЩЕНИЯ
Введение
Подвижность и перемещение живых структур имеет много общих черт. В их основе лежат сходные по своей структуре и функции сократительные белки актина и миозина, взаимодействие которых, в принципе, осуществляют необходимые перемещения в пространстве. Существуют специализированные структуры, выполняющие сократительные функции – мышечная ткань, хотя сократительные белки (актин) являются обязательным компонентом цитоскелета любой клетки.
Виды мышечного сокращения: Изометрическое – при неизменных размерах Изотоническое – при неизменной силе.
Ауксотоническое – при изменении длины и силы.
Мышечная ткань подразделяется на:
1.Поперечно-полосатую (скелетная и сердечная) мускулатуру.
2.Гладкие мышцы.
Их отличия касаются структуры, функции и эмбриогенеза.
Скелетные мышцы
Структурно-функциональной единицей скелетной мышцы является
мышечное волокно – многоядерная клетка, образовавшаяся из мезодермы слиянием нескольких сотен, а иногда и тысяч клеток. Длина мышечного волокна достигает 100-1000мкм., толщина 20-80 мкм Покрыто мышечное волокно двухслойной мембраной – сарколеммой. Непосредственно к цитоплазме прилежит плазмалемма – обычная плазматическая мембрана толщиной 7,5нм. Через некоторое пространство, заполненное соединительнотканными волокнами, располагается базальная мембрана (5,5нм), которая является общей для пучка из 20-40 мышечных волокон.
Основное внутриклеточное пространство мышечного волокна заполнено миофибриллами диаметром 1-2 мкм. Их около 2000 и они тянутся вдоль всего мышечного волокна и сформированы в саркомеры.
Саркомер скелетных мышц. Особенностью скелетной, как и всех поперечно-полосатых мышц, является поперечная исчерченность внутреннего содержимого цитоплазмы характерными структурами саркомерами (
Рис. 28).
127

Рис. 28. Саркомер скелетных
мышц
Длина саркомера (от Z- пластины до Z-пластины) в скелетных мышцах составляет 2,2мкм. В этих структурах располагаются основные сократительные белки актин и миозин. Строгая упорядоченность их в саркомерах приводит с чередованию оптически более плотных и менее плотных структур.
К Z-пластинам саркомера симметрично по обе стороны прикрепляются нити актина. Между ними в оптически менее плотной (изотропной) зоне I-дисков расположены нити миозина. Посредине каждого I-диска имеется М-полоса – особая мембрана, на которой фиксируются нити миозина. Частично нити актина и миозина перекрываются, образуя оптически более плотную (анизотропную) зону или А-диск. Светлую часть А-диска Н-полосу, содержащую только нити актина, посредине пересекает Z-пластина.
Триада скелетных мышц представляет собой совокупность структур, обеспечивающих запуск сокращения в ответ на раздражение сарколеммы. Она образована тремя структурами (см.Рис):
1.Т-системой – впячивания плазматической мембраны внутрь мышечного волокна диаметром около 0,03 мкм.
2.Концевыми цистернами саркоплазматического ретикулума (СПР).
3.Продольными каналами СПР.
Обычно триада располагаются вблизи Z-пластин саркомера.
Структура и функция сократительных белков
Основную сократительную функцию во всех видах мышц осуществляют тонкие и толстые нити-миофиламенты (миофибриллы) актин и миозин.
Вспомогательную – регуляторную осуществляют тропомиозин (TrM, ММ:68 кD) и комплекс тропонина (Tr, ММ:70 кD), который состоит из субединиц (Рис. 29)
128

Рис. 29. Структура сократительных белков
1.Ингибиторный тропонин – TrI, ММ:21 kD;
2.Тропонинсвязывающий тропонин – TrT ММ:31 kD;
3.Кальций-связывающий тропонин – TrC, ММ:18 kD.
Молекула актина встречается в двух формах: глобулярной (G) и
фибриллярной (F). Переход из G- в F-форму происходит в присутствии АТФ и ионов кальция непосредственно перед сокращением. G-форма представляет собой глобулу ММ 48 кD. Полимеризованная F-форма собирается на тропомиозине вместе с тропониновым комплексом: на одной молекуле TrM+ Tr(I,T,C) обнаруживается 7 G-глобул актина. Кроме того, в окончательном варианте две таких структуры закручиваются относительно друг друга, образуя суперспираль – миофибриллу актина, непосредственного участника сокращения.
Молекула миозина (ММ:480 кD) состоит из 2-х тяжелых (2x200 кD) и 4-х легких цепей. Две из них (18,5 кD) –отщепляются при обработке миозина 5,5- дитиобис-2-нитрбензойной кислотой (ДТНБ цепи). Две другие (20,7 кD и 16,5 kD) диссоциируют в щелочной среде и названы щелочными. Легкие цепи формируют головку молекулы миозина, обладающей способностью к АТФ-азной активности (щелочные цепи) и изменению конформации под воздействием ионов кальция (ДТНБ-цепи). Толстая миофибрила содержит около сотни молекул миозина, закрученных относительно друг друга тяжелыми цепями от центра к краям. В результате, центральная область
129

толстых нитей миозина не содержит головок легких цепей, обладающих АТФазной активностью.
Молекулярные механизмы мышечного сокращения
Для осуществления сокращения мышечного волокна требуется ряд условий:
1.Развитие потенциала действия на сарколемме.
2.Повышение внутриклеточной концентрации ионов кальция выше 0,1
мкМ.
3.Наличие в цитозоле молекул АТФ.
Потенциал действия (ПД) скелетного волокна имеет Na+,K+-ю природу (
Рис. 30).
Рис. 30. Электрическая и
сократительная активность мышечного волокна
Его амплитуда достигает 120-130 мВ, овершут +30-+50 мВ, продолжительность 3-5 мс. Особенностью является длительная следовая деполяризация (около 10 мс), обусловленная аккумуляцией ионов K+ в Т- трубочках. Их разрушение при действии глицерина удаляет не только следовый потенциал, но и делает невозможным развитие сокращение на фоне ПД.
Повышение внутриклеточной концентрации ионов кальция происходит в результате его высвобождения из концевых цистерн СПР после развития ПД.
Взаимодействие электрических процессов на сарколемме (ПД) и непосредственно сократительных белков осуществляется с помощью процесса электромеханического сопряжения (ЭМС). В настоящее время существует несколько гипотез развития ЭМС:
1.Морфологическая, которая предполагает наличие структур, связывающих Т-систему и СПР.
2.Электрическая, которая предполагает перераспределение электрического поля с Т-системы на СПР.
3.Химическая, которая предполагает, что в процессе развития ПД внутрь волокна выделяется вещество-посредник запуска сокращения, например, инозитолтрифосфат.
После повышения внутриклеточного уровня выше критического (0,1 мкМ) ионы кальция соединяются с TrС, в результате чего весь тропониновый
130