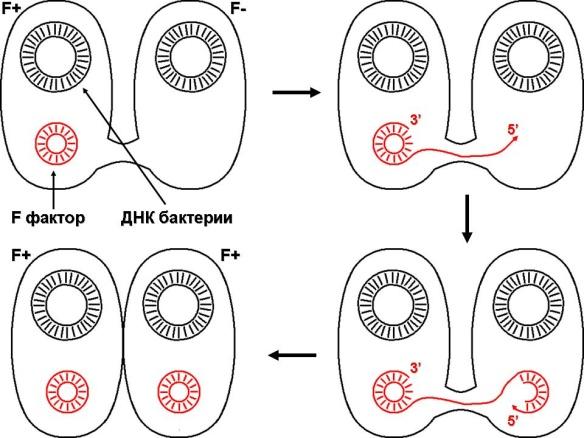
1.42. Фотография «Конъюгация у бактерий под электронным микроскопом»
1 - процесс конъюгации между F+ и F- бактериями
2 - на электронной фотографии - клетки донора (F+) и реципиента (F-), хорошо виден цитоплазматический мостик;
3 - передача генетического материала в редких случаях может происходить по половым пилям, но все-таки в подавляющем большинстве случаев она происходит по цитоплазматичеекому мостику
4 - существуют два состояния полового фактора: свободное нахождение в цитоплазме, и тогда бактерии обозначаются F* и интегрированное в хромосому. В последнем случае бактерии обозначаются Hfr, т.е.обладающис высокой частотой рекомбинации (от англ.: hight frequence of recombinantion). Если половой фактор находится в свободном состоянии в цитоплазме, то в клетку реципиент передается только его генетический материал.
5 - передача генов резистентности к антибиотикам, кодирующих синтез ферментов, факторов вирулентности, ускорение процесса репродукции у бактерий в случае формирования Hfr-гклеток
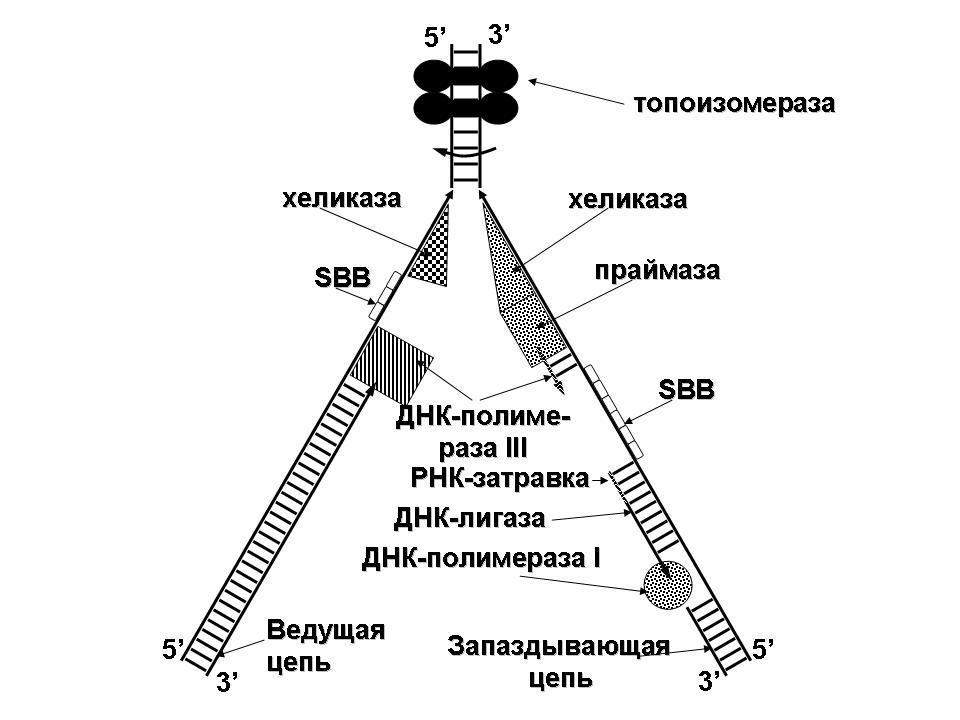
1.43. Схема «Формирование репликативной вилки хромосомы»
1 - схема репликации ДНК (формирование репликативной вилки)
2 - процесс расплетания нитей ДНК и участвующие в нём ферменты
3 - синтез ДНК на обеих матричных цепях идет в направлении от 5' к 3' концам. На ведущей нити ДНК он происходит непрерывно, а на отстающей прерывисто в виде фрагментов Оказаки, каждый из которых включает 1000-2000 нуклеотидов
4 - образование репликативной вилки ДНК начинается в определённом участке ДНК (ориджин) при участии фермента ДНК-гиразы; дальнейшее формирование репликативной вилки обеспечивают ферменты хелнказы. Одна хеликаза передвигается по ведущей нити ДНК, а другая по отстающей. Кроме хеликаз в формировании репликативной вилки участвуют SSB - белки, специфически связывающиеся с каждой из расплетенных нитей ДНК. Они предотвращают преждевременное их соединение. Вновь синтезированные нити ДНК всегда содержат на 51 концах несколько рибонуклеотидов, т.е. Синтез ДНК начинается с синтеза короткой цепи РНК, связанной с матрицей ДНК. Эту РНК-затравку образует специальный фермент, который называется ДНК-праймаза (праймер - затравка). Затем синтез ДНК продолжается благодаря присоединению к РНК-затравке дезокеирибонуклеотидов с помощью ДНК-полимиразы III. Удаление РНК-затравки и застройка брешей осуществляется ДНК-полимеразой I. На отстающей нити ДНК, остающиеся между двумя соседние фрагментами Оказаки, однонитевые разрывы заделывает ДНК-лигаза. Раскручивание узлов, образующихся перед репликативной вилкой сверхсперилизованными нитями ДНК, осуществляет фермент топоизомераза. Энергия, необходимая для расплетания нитей ДНК поставляется в результате гидролиза АТФ
5 - делению бактериальной клетки обязательно должна предшествовать репликация ДНК
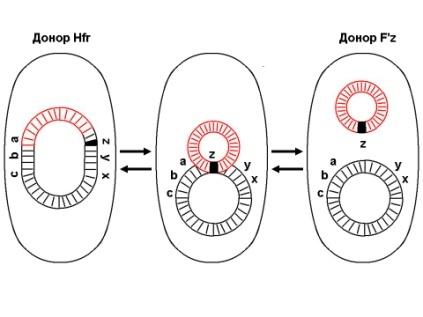
1.44. Схема «Конъюгация у бактерий. Формирование f рекомбинантов»
1 - конъюгация у бактерий: формирование Ррекомбинантов
2 - донор Hfr, вычленение плазмиды с геном z, донор f'z
3 - существуют два состояния полового фактора: свободное нахождение в цитоплазме, и тогда бактерии обозначаются F*" и интегрированное в хромосому - обозначаются Hfr, т.е.обладающие высокой частотой рекомбинации (от англ.: hight frequence of recombinantion). Бели половой фактор находится в свободном состоянии в цитоплазме, то в клетку реципиент передается только его генетический материал
4 - У Hfr бактерий может происходить выход полового фактора из состава хромосомы в цитоплазму, в результате возникает клон F* клеток. В некоторых случаях при выходе F фактора он может включить расположенный рядом ген бактериальной хромосомы (путем рекомбинации). В таких случаях рядом с обозначением полового фактора дается символ соответствующего гена (z). Следовательно, особенностью Hfr бактерий является передача генов собственной хромосомы в клетку реципиент. Например, fmac, т.е. В составе полового фактора находится лактозный оперон, отвечающий за утилизацию лактозы. В результате бактерии имеют делецию, соответствующую лактозному гену, являющегося уже составной частью полового фактора. Если в состав полового фактора войдут гены, жизненно необходимые бактерии, то потеря его клеткой может привести к ее гибели. Поскольку половые факторы могут включаться в различные участки бактериальной хромо сомы, то F плазмида может содержать различные гены.
5 - передача генов резистентности к антибиотикам, кодирующих синтез ферментов, факторов вирулентности, ускорение процесса репродукции у бактерий в случае формирования Hfr-гклеток