

1.1Анатомия вестибулярного анализатора
Вестибулярный анализатор - парный орган равновесия, состоящий из периферического и центрального отделов. У человека помимо вестибулярного аппарата равновесие зависит от взаимодействия органов слуха, зрения, от функции мозжечка и коры головного мозга, от проприоцептивных и тактильных раздражений, идущих, например, от чувствительных к давлению рецепторов в подошвах стоп.
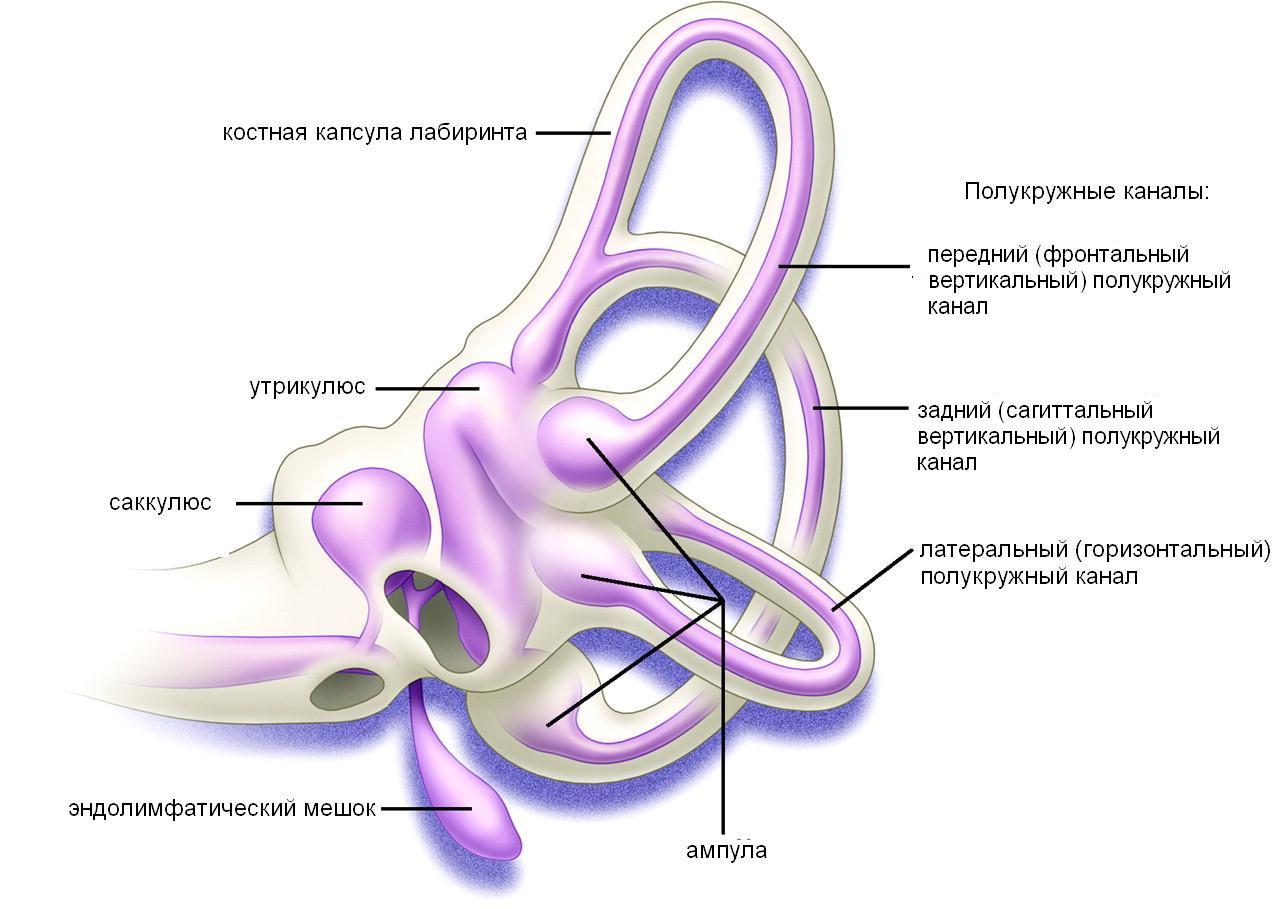
Рассмотрим вначале строение периферических вестибулярных рецепторов, находящихся в вестибулярном отделе перепончатого лабиринта. Он представляет собой систему сложных полостей: трех полукружных каналов и преддверия, с которым все эти каналы соединены своими концами. В каждом полукружном канале в свою очередь заключены мембранные протоки. В преддверии находятся два мембранозных мешочка – сферический (саккулюс) и элиптический (утрикулюс). Мембранозные структуры (и каналы и мешочки) заполнены эндолимфой и окружены перилимфой, которая заполняет костные каналы и преддверие. Мембранозные протоки связаны многочисленными соединительнотканными трабекулами с костными стенками, а одной своей стороной они прилежат к стенке костного канала.
Преддверие,центральная часть лабиринта, филогенетически наиболее древнее образование внутреннего уха, представляющее собой небольшую полость, внутри которой различают два кармана: сферический (recessussphericus) и эллиптический (recessusellipticus). В первом, расположенном возле улитки,сферический мешочек(sacculus). Во втором кармане, примыкающем к полукружным каналам -эллиптический мешочек(utriculus). На наружной стенке преддверия имеется окно преддверия, обращенное в полость среднего уха и прикрытое с его стороны основанием стремени. Передняя часть преддверия (сферическая) через лестницу преддверия сообщается с улиткой, задняя (эллиптическая) -с полукружными каналами.
В саккулюсе и утрикулюсе помимо эндолимфы имеются рецепторные образования, называемые пятнами (maculaе), которые являются рецепторами пространственного чувства. Эти рецепторные образования называются отолитовыми рецепторами и располагаются на небольших возвышениях внутренней поверхности обоих мешочков и соответственно мешочкам называютсяmaculautriculiиmaculasacculi. Отолитовые рецепторы (от названия входящих в их состав специфических анатомических образований, рассматриваемых далее), как и ампулярные, также ориентированы во взаимно перпендикулярных плоскостях: в эллиптическом мешочке рецептор расположен на ее дне горизонтально, а в находящемся возле улитки сферическом - на медиальной стенке мешочка вертикально.
Отолитовый рецептор тоже состоит из опорных и волосковых клеток. Длинные волоски чувствительных клеток, переплетаясь своими концами, также образуют сеть, которая погружена в желеобразную массу. Но в отличие от ампулярного рецептора желеобразная субстанция отолитового аппарата содержит большое число кристаллов, имеющих форму параллелепипедов. Кристаллы эти называются отолитами(от греч.otos-yхо,litos- камень) и состоят из фосфата и карбоната кальция (аррагонит). Длинные волоски рецепторных клеток вместе с отолитами и желеобразной массой образуютотолитовую мембрану.Благодаря содержанию отолитов эта мембрана обладает большой удельной массой и легко смещается даже при небольших передвижениях головы. Раздражение отолитовых рецепторов давлением или смещением отолитовой мембраны при прямолинейных ускорениях либо воздействии силы тяжести приводят к трансформации механической энергии в электрическую. Эта энергия передается напериферические отолитовые отросткивестибулярного ганглия.
Оба мешочка соединены между собой посредством тонкого канала (ductusutriculosacuilaris), который имеет ответвление -эндолимфатический проток (ductusendolymphaticus). Этот проток проходит в костном канале, или водопроводе преддверия (aqueductusvestibuli), а затем на задней поверхности пирамиды височной кости слепо заканчивается расширением – эндолимфатическим мешком (saccusendolymphaticus) в дупликатуре твердой мозговой оболочки на задней поверхности пирамиды височной кости.
Полукружные каналы.Во внутреннем ухе с каждой стороны различают три полукружных канала. Диаметр костных полукружных каналов около 1.5 мм, длина – от 12 до 18 мм. Они находятся в трех взаимно перпендикулярных плоскостях: один - в горизонтальной и два - в вертикальной. Однако ориентация каналов не совсем совпадает с пространственной троичной системой координат. Поэтому, в какой бы плоскости ни происходило движение головы, раздражение вестибулярных рецепторов будет осуществляться во всех каналах, но больше в том, который находится в плоскости вращения. Наружный (canalissemicircularislateralis) или горизонтальный полукружный канал располагается под углом 30°к горизонтальной плоскости. Передний (canalissemicircularisanterior), или фронтальный вертикальный, находится под углом 450 к фронтальной плоскости. Задний полукружный канал (canalissemicircularisposterior), или сагиттальный вертикальный - под углом 450 к сагиттальной плоскости. Каналы расположены таким образом, что каждый из них лежит в плоскости, перпендикулярной плоскостям двух других каналов, отсюда и их название. Латеральный канал контролирует горизонтальную плоскость, передний вертикальный контролирует фронтальную плоскость, а задний контролирует сагиттальную плоскость. Вертикальные каналы ориентированы так, что передний фронтальный канал одного лабиринта лежит в одной плоскости с задним сагиттальным каналом противоположного уха.
В каждом канале имеются два колена (ножки): простое - гладкое и грушевидно расширенное -ампулярное. Гладкие ножки верхнего и заднего вертикальных каналов слиты в одну общую ножку (cruscommune), а горизонтальный полукружный канал имеет собственную гладкую ножку. Все пять ножек полукружных каналов (две гладкие и три расширенные) обращены к заднему эллиптическому карману преддверия.
Перепончатые полукружные протоки и мешочки преддверия. Перепончатые полукружные протоки расположены в костных каналах. Они меньше по диаметру и повторяют их строение, т.е. имеют расширенные ампулярные и простые гладкие части (ножки). Одной из своих соединительнотканных стенок перепончатый полукружный проток прилежит к стенке костной капсулы, оставляя значительное место для перилимфатического пространства. Исключение составляют ампулы перепончатых каналов, которые почти полностью заполняют просвет костных ампул. Внутренняя поверхность перепончатых протоков выстлана эндотелием, за исключением ампул, в которых расположены рецепторные клетки. На внутренней поверхности ампул имеется круговой выступ -гребень (cristaampullaris), который состоит из двух слоев клеток -опорных и чувствительных волосковых. Волосковые клетки являются периферическими рецепторами вестибулярного нерва.
У человека, как и у других высших позвоночных, вестибулярные рецепторы внутреннего уха представлены чувствительными к раздражению грушевидными механорецепторами Iтипа. Филогенетически они моложе менее чувствительных цилиндрических клетокIIтипа, появляющихся у круглоротых рыб и амфибий. Для клетокIтипа характерной особенностью является то, что нервное окончание периферического отростка вестибулярного ганглия образует объемное ложе («чашу»), которое почти полностью вмещает в себя тело клетки. Благодаря такому обширному синаптическому контакту чувствительной рецепторной клетки, а также тому, что афферентные нервные волокна, подходящие к клетке, являются одними из самых толстых (до 20 мкм), обеспечивается высокая надежность функции вестибулярных рецепторов.
Каждую волосковую клетку венчает один длинный и толстый волосок – киноцилий (cino– подвижный,cilia– волосок), располагающийся с краю верхушки клетки. Остальную ее поверхность покрывают от 50 до 110 более тонких и коротких волосков –стереоцилий(stereo– твердый, неподвижный). Между киноцилией и стереоцилиями существует разность потенциалов, которая зависит от их взаимного расположения. Когда киноцилия приближается к стереоцилиям или, напротив, удаляется от них, разность потенциалов уменьшается или возрастает (происходит гипо- или гиперполяризация). Это сопровождается соответственно снижением или возрастанием импульсации рецептора, постоянно находящегося в активном состоянии. Длинные волоски нейроэпителиальных клеток (киноцилии) склеены между собой эластичной желеобразной массой, состоящей из сульфомукополисахаридов, и наcristaampullarisсоставляют образование в виде круговой заслонки-кисточки (cupulaterminalis). Эта кисточка-купула, как поршень, почти полностью закрывает просвет ампулы, поэтому любое перемещение эндолимфы в перепончатом канале вызывает отклонение купулы. Возникающее при угловых движениях головы (повороты, наклоны) механическое передвижение эндолимфы смещаетcupulaterminalisв сторону ампулы или гладкого колена перепончатого канала. Механическая энергия смещения купулы является раздражителем для нейроэпителиальных волосковых клеток. Эта энергия преобразуется в волосковых клетках в электрический импульс и передается напериферические ампулярные отросткилежащего в глубине внутреннего слухового проходавестибулярного ганглия(ganglionvestibularisScarpae).
Таким образом, вестибулярные сенсорные клетки каждого внутреннего уха расположены в пяти рецепторных областях: по одной в ампулах трех полукружных протоков и по одной в двух мешочках преддверия. Соответственно пространственной ориентации этих вестибулярных рецепторов наш орган равновесия получает исчерпывающую информацию о положении и движении тела в любой плоскости и направлении. К чувствительным клеткам ампулярных и отолитовых рецепторов подходят два отдельных пучка периферических волокон (аксонов) от биполярных клеток вестибулярного ганглия (gangl.Scarpаe), располагающегося во внутреннем слуховом проходе (I нейрон). Верхний периферический пучок идет к чувствительным волосковым клеткам элиптического мешочка, а также к ампулярным рецепторам латерального и переднего (верхнего) полукружных протоков. Нижний периферический пучок подходит к заднему полукружному протоку и сферическому мешочку. Центральные волокна биполярных клеток вестибулярного ганглия образуютвестибулярный нерв (radixvestybularis) и идут по внутреннему слуховому проходу височной кости в одной нервной оболочке со слуховым нервом, формируяVIIIчерепной мозговой нерв (n.cochleovestybularis), к мостомозжечковому углу соответствующей стороны. Вместе сVIIIчерепным мозговым нервом во внутреннем слуховом проходе следует и лицевой нерв (n.facialis,VIIчерепной нерв), что имеет важное значение в отоневрологической диагностике периферических и центральных вестибулярных нарушений.
После выхода из внутреннего слухового прохода височной кости вестибулярный и слуховой нервы на уровне мостомозжечкового угла проникают в ствол мозга, где в продолговатом мозге вестибулярный нерв передает по восходящим вестибулярным волокнам (числом до 12 000) возбуждение на вестибулярные ядра, а по нисходящим (числом не более 200) – получает обратную регулирующую информацию от ядер ретикулярной формации продолговатого мозга и некоторых других мозговых структур. Вспомним, что ретикулярная формация мозга является своеобразным тормозящим и активирующим центром различных подкорковых отделов ЦНС. Она регулирует, например режим сна и бодрствования, отвечает за процессы внимания и тренировки. В мосту восходящие волокна вестибулярной порцииVIIIнерва снова делятся на восходящие и нисходящие пучки и направляются к вестибулярным ядрам своей стороны.
На границе между продолговатым мозгом и стволом, на дне IVжелудочка, по обе стороны ромбовидной ямки, в латеральных ее углах располагаются правый и левый ядерные вестибулярные комплексы (II нейрон). В вестибулярном комплексе соответствующей стороны находятся четыре вестибулярных ядра.Верхнее,nucl.vestibularissuperior, имедиальное ядра,nucl.vestibularismedialis, получают афферентные волокна от ампулярных рецепторов и из мозжечка.Нижнее ядро,nucl.vestibularisinferior, возбуждается, в основном, от отолитовых и частично от ампулярных рецепторов внутреннего уха.Латеральное ядро,nucl.vestibularislateralis, принимает импульсы от мозжечка и отолитовых рецепторов. Восходящие пучки вестибулярного корешкаVIIIнерва заканчиваются синапсами в клетках верхнего вестибулярного ядра. Нисходящие – в клетках остальных трех ядер.
Далее аксоны клеток вестибулярных ядер формируют несколько ассоциативных (связующих) пучков с другими отделами ЦНС, или трактов, благодаря которым осуществляется полноценная пространственная ориентация организма, а также обеспечиваются высокие приспособительные и компенсаторные возможности вестибулярной системы.
С позиций клинической анатомии и дифференциальной диагностики вестибулярных расстройств следует выделить пять таких трактов, которые связывают вестибулярные ядра с последующими нейронами соответствующих рефлекторных дуг.
1). Вестибулоспинальный тракт, или пучок,tr.vestibulospinalis, начинается от латеральных ядер (Дейтерса) по обе стороны ромбовидной ямки и посегментно заканчивается в двигательных промежуточных нейронах передних рогов спинного мозга. Промежуточные мотонейроны возбуждают моторные альфа- и гамма- нейроны мышц-разгибателей, т. е. они являются антагонистами пирамидного пути и осуществляют угнетение мышц-сгибателей и возбуждение мышц-разгибателей. Этот механизм представляет собой часть статокинетической системы, служащей для поддержания равновесия. Вестибулоспинальный тракт в содружестве с ретикулоспинальным и медиальным продольным трактами обеспечивает безусловнорефлекторное поддержание равновесия тела за счет перераспределения тонуса мышц шеи, конечностей и туловища.
В безусловнорефлекторной регуляции мышечного тонуса принимает участие и экстрапирамидная система: часть аксонов клеток от латерального и медиального вестибулярных ядер формируют вестибулоталамический тракт, tr.vestybulothalamicus. Из курса неврологии вспомним, что пирамидная система контролирует произвольные мышечные движения и проводит программу таких движений от коры больших полушарий до ствола головного и спинного мозга. В частности, она включает в себя двигательные ядра черепных нервов и серого вещества спинного мозга. Экстрапирамидная система осуществляет регуляцию непроизвольных компонентов моторики, как безусловных, так и условнорефлекторных (профессиональные навыки). Она объединяет двигательные центры коры головного мозга, его ядра и проводящие пути, которые не проходят через пирамиды продолговатого мозга. Помимо других образований, в нее входят ядра таламуса, мозжечок, ретикулярная формация. Экстрапирамидная система управляет мышечным тонусом, координацией движений, позной устойчивостью, создает фон для реализации быстрых, точных и дифференцированных движений. Обе системы активно взаимодействуют: экстрапирамидная система обеспечивает упорядоченный ход произвольных движений, регулируемых пирамидной системой.
2). Медиальный продольный тракт, или пучок,tr.vestybulolongitudinalis, связывает вестибулярные ядра (верхнее, нижнее, медиальное) с ядрами трех глазодвигательных нервов: глазодвигательного (III), блокового (IV) и отводящего (VIчерепной нерв). По этому тракту осуществляются связи, обеспечивающие компенсаторные рефлекторные повороты глаз во время движений головы. Особая роль в координации движений глаз и головы принадлежит медиальному ядру, так как посредством своих проекций в медиальном продольном пучке это ядро согласовывает глазодвигательные и шейные моторные реакции. На изучении вестибулярно обусловленных движений глаз основано большинство методик экспериментальной и клинической вестибулологии. Эта форма подсознательных движений глазных яблок называетсямедленным компонентом вестибулярного нистагма, иливестибулоокулярным рефлексом(ВОР). Основной функцией ВОР является стабилизация положения глазных яблок по отношению к голове во время ходьбы, бега, прыжков, поворотов и других движений. Благодаря этому изображение окружающего пространства на сетчатке глаз остается относительно неподвижным.
Вестибулоокулярный рефлекс является важным компонентом в группе глазодвигательных рефлексов. Он помогает сохранять достаточную остроту зрения в динамических условиях, т.е. участвует в поддержании динамического зрения. На глазодвигательные ядра оказывают дополнительное влияние и обширные мультисинаптические связи от ретикулярной формации. Нижняя часть медиального продольного пучка также обеспечивает связь с моторным отделом спинного мозга.
3). Вестибуломозжечковый тракт,tr.vestibulocerebellaris, имеет афферентный и эфферентный отделы. Афферентный вестибуломозжечковый тракт проходит от вестибулярных ядер (верхнее, медиальное и латеральное) через нижние ножки мозжечка и заканчивается в коре червя мозжечка. Мозжечок является высшим безусловнорефлекторным центром двигательной координации.
От мозжечка импульс возвращается, в частности, к вестибулярному анализатору по эфферентному вестибуломозжечковому пути и при помощи латерального ядра оказывает коррегирующее мозжечковое влияние на спинной мозг, а посредством верхнего и медиального вестибулярных ядер - на глазодвигательные центры.
4). Часть аксонов клеток латерального ядра вступает в состав заднего продольного пучка (тракта).Этот тракт соединяет задние гипоталамические ядра, являющиеся центрами мозговой интеграции вегетативных процессов, с вегетативными ядрами глазодвигательного (III), лицевого (VII), языкоглоточного (IX) и блуждающего (X) нервов, а также с вегетативными симпатическими и парасимпатическими центрами спинного мозга. Задний продольный пучок способствует реализации вегетативных реакций, связанных с работой внутренних органов. Поэтому, например, при вестибулярном раздражении изменяется сосудистый тонус, учащаются сердечный ритм и дыхание, усиливается перистальтика кишечника. Гуморальные вестибулярные реакции гипоталамо-гипофизарно-надпочечниковой системы запускаются благодаря вестибуло-таламо-гипоталамическим связям.
5). Вестибулокортикальный тракт,tr.vestibulocorticalis, или вестибулярный сенсорный тракт, обеспечивается системой вертикальных волокон, идущих от всех четырех ядер. Этот тракт переключается в таламусе (III нейрон), являющемся подкорковым сенсорным центром, направляется к задней ножке внутренней капсулы ( подкорковое образование, через которое проходят все проекционные волокна различных зон коры больших полушарий) и оканчивается на клетках височно-теменной области (средняя и нижняя извилины) коры головного мозга (IV нейрон) со своей стороны. Следует подчеркнуть, что четкого коркового представительства у вестибулярного анализатора нет. Рассеянные по соответствующей области мозговой коры клетки вестибулярного анализатора имеют обширные связи с соседними сенсорными корковыми центрами: зрительным (затылочная доля), слуховым (верхняя височная извилина), координаторным проприоцептивным (передняя и задняя центральные извилины и верхняя теменная долька – центр схемы тела) и другими.
Ощущение головокружения при вестибулярной надпороговой стимуляции возникает и передается по этому тракту. Адаптационные реакции обеспечиваются связями вестибулярной системы с гипоталамусом посредством ретикулярной формации и через нее же – с корой головного мозга, благодаря чему формируется осознанное восприятие движения и положения тела в окружающем пространстве.
Таким образом, многочисленные афферентные пути, идут от вестибулярных ядер к мозжечку, ретикулярной формации, спинному мозгу и противоположным вестибулярным ядрам. Важно, что от одного вестибулярного ядра отходит несколько различных ассоциативных путей. Это нужно учитывать как в оценке нормы вестибулярной функции, так и при выявлении вестибулярной патологии различного происхождения.
Реакции, возникающие при раздражении вестибулярных рецепторов патологическим процессом или вызываемые искусственно, распространяются по всем его связям и вызывают соответствующее рефлекторное действие. Все вестибулярные реакции условно объединяют в три группы: вестибулосоматические (двигательные), вестибулосенсорные и вестибуловегетативные.
Вестибулосоматические реакции опосредованы через вестибулоспинальный, вестибулоглазодвигательный и вестибуломозжечковый тракты. Обусловленные вестибулярным раздражением сенсорные реакции опосредованы через вестибулокортикальный тракт. Через задний продольный пучок и ретикулярную формацию реализуются вестибуловегетативные реакции. Наиболее информативным является вестибулярный рефлекс на глазодвигательные ядра, вызывающий нистагм.