

2. Особенности вирусНой инфекцИи
По основным признакам вирусные инфекции не отличаются от инфекций бактериальной или другой этиологии. Выделяют те же периоды инфекционного процесса (инкубационный, продромальный, основных клинических проявлений, исхода). Исходы вирусных инфекций те же: реконвалесценция, летальный исход, хронизация процесса, носительство.
Вирусные инфекции имеют те же резервуары и источники - человек, животное (больные и носители), за исключением объектов внешней среды, среди вирусных инфекций нет сапронозов.
При вирусных инфекциях те же пути передачи (воздушно-капельный, фекально-оральный, контактный, трансмиссивный и инъекционный, трансплацентарный и др.), те же входные ворота и пути распространения по организму и выделения из него.
При вирусных инфекциях так же развиваются иммунологические сдвиги в организме, остается иммунитет, иногда пожизненный.
Таким образом, вирусы выступают как типичные возбудители инфекционных заболеваний.
Самое существенное отличие вирусных инфекций от бактериальных с точки зрения практического здравоохранения это недостаточность терапии, отсутствие до настоящего времени эффективных и безвредных средств лечения. Имеющиеся в последнее время сдвиги в этом направлении пока не меняют в целом этого положения. Антибиотикотерапия при вирусных инфекциях как этиотропная терапия неэффективна.
Профилактика же вирусных инфекций аналогична профилактике бактериальных, неспецифические методы профилактики, направленные на разрыв эпидемической цепи, те же самые, специфические методы профилактики основываются на применении вакцин и сывороточных препаратов.
Подчеркивают еще то, что существуют сборные группы вирусов, вызывающих массовые инфекционные заболевания: респираторные, гастроэнтериты, гепатиты. Но это несущественное отличие, так как можно выделить и группы бактериальных массовых инфекций, просто мы умеем с ними лучше бороться, чем с вирусными, поэтому массовые бактериальные инфекции менее заметны для медицины.
Вместе с тем, имеются достаточно существенные отличия вирусных инфекций от бактериальных.
Взаимодействие вируса и хозяина может рассматриваться на различных уровнях - на уровне клетки, организма и популяции или общества.
На клеточном уровне вирусная инфекция может вызвать очень широкий диапазон эффектов, от отсутствия видимых клеточных повреждений, до быстрого разрушения клеток. Некоторые вирусы (например, полиовирус, возбудитель полиомиелита) вызывают гибель клетки (цитоцидный эффект) или даже лизис (цитолиз). Другие могут вызывать пролиферацию (размножение) клеток, например, возбудитель контагиозного моллюска, или злокачественную трансформацию (например, онкогенные вирусы). В некоторых случаях вирус и клетка - хозяин вступает в мирное сосуществование, оба размножаются независимо друг от друга без какого-либо ущерба для клетки, состояние, известное как «стационарная инфекция».
В культуре тканей вирусная инфекция может приводить к легко заметным изменениям клеток (цитопатическое действиe вирусов, ЦПД). Они могут не быть аналогичны изменениям у инфицированного животного, поскольку в этом случае инфекция находится под влиянием различных механизмов защиты организма.
Поражения клеток могут развиваться по многим причинам. Ранние или неструктурные вирусные белки часто вызывают остановку синтеза белка и ДНК хозяина. Большие количества вирусных макромолекул, которые накапливаются в инфицированной клетке, могут нарушать клеточную архитектуру и вызывать токсическое действие. Может изменяться проницаемость плазматических мембран с выходом лизосомальных ферментов, что приводит к автолизу клеток.
Многие вирусы вызывают изменения в цитоплазматической мембране инфицированных клеток. Некоторые (например, респираторно-синцитиальный вирус) вызывают слияние смежных клеточных мембран, приводя к формированию поликариоцитоза (многоядерности) или синцития. Вирусиндуцированные антигены могут появляться на поверхности инфицированных клеток. Эти антигены могут придавать клеткам новые свойства. Например, вирусный гемагглютинин появляется на поверхности клеток, инфицированных вирусом гриппа, и вызывает адсорбцию эритроцитов на поверхности клетки (гемадсорбцию). Вирусиндуцированные антигены могут также появляться на поверхности клеток, трансформированных онкогенными вирусами.
Некоторые вирусы, например, кори, паротита, цитомегалии, ветряной оспы, аденовирусы вызывают повреждения хромосом клеток хозяина. В клетках, инфицированных аденовирусами типа 12 и 31, часто наблюдаются пробелы и разрывы хроматид.
Наиболее характерной гистологической особенностью инфицированной вирусом – появление телец включений. Тельца-включения - структуры различных размеров, формы, местоположения и способностью к окраске, которые могут обнаруживаться в инфицированных вирусом клетках под оптическим микроскопом. Они могут быть расположены в цитоплазме (например, включения поксвирусов), ядре (например, вирусов герпеса) или и там и там (например, вируса кори). Они обычно ацидофильны и могут быть замечены как розовые структуры при окраске по Романовскому-Гимзе или эозин-метиленовым синим. Некоторые вирусы (например, аденовирус) формируют базофильные включения.
Выявление телец-включений помогает в диагностике некоторых вирусных инфекций. Присутствие внутриклеточных эозинофильных включений (телец Негри) в мозговых клетках животных подтверждает предполагаемый диагноз бешенства. В клетках, инфицированных вирусом коровьей оспы, обнаруживаются многочисленные довольно мелкие включения, известные под названием телец Гварниери. Внутриядерные тела включения классифицируются на два типа. Включения типа А имеют различный размер и зернистый вид, пример, у вирусов герпеса, желтой лихорадки), в то время как включения типа B более очерчены и часто множественны (например, у аденовируса).
Тельца-включения могут быть кристаллическими агрегатами вирионов или состоять из вирусных антигенов, присутствующих в месте синтеза вируса. Некоторые включения представляют дегенеративные изменения, вызванные вирусной инфекцией, которые придают клетке способность к измененной окраске.
Отметим, что вирусная инфекция на клеточном уровне может быть автономная (продуктивная и абортивная) и интегративная (с неопластической трансформацией и без неё).
При автономной инфекции вирусный геном реплицируется независимо от клеточного генома, между ними нет физической связи, хотя они и взаимодействуют в ходе инфекции. Продуктивная инфекция завершается образованием инфекционного потомства вирусов, абортивная инфекция (прерванная) может не завершаться образованием инфекционных вирионов или же они образуются в гораздо меньшем количестве, чем при продуктивной инфекции. Абортивная инфекция возникает при заражении клеток дефектным вирусом, при неблагоприятных условиях (повышение температуры, изменение рН в очаге воспаления, наличии вирусных ингибиторов). Дефектные вирусы не имеют полного генома и требуют присутствия вируса-помощника (аденоассоциированный парвовирус, дельта-фактор при гепатите В).
Кроме того, в популяции вируса при его репродукции вместе с инфекционными вирионами могут накапливаться и так называемые ДИ-частицы (дефектные интерферирующие частицы), они могут давать лишь абортивную инфекцию, так как лишены части генома. В то же время, наличие таких дефектных частиц обеспечивает одну из форм длительного нахождения вируса в клетке - персистенцию.
В основу классификации вирусных инфекций на уровне организма положены признаки: степень генерализации, продолжительность инфекции, клиническое течение, выделение вируса из организма.
|
|
генерализованная инфекция | ||||||||||
|
|
персистентная
|
острая |
персистентная | ||||||||
|
явная |
скрытая |
|
латентная |
хроническая
|
|
явная |
скрытая |
|
латентная |
хроническая |
медленная |



 очаговая
инфекция
очаговая
инфекция






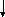
 острая
острая
При очаговой инфекции действие вируса проявляется непосредственно во входных воротах в связи с его локальной репродукцией, при генерализованной инфекции после ограниченного периода репродукции вируса во входных воротах происходит раcпространение вируса по организму генерализация процесса. Очаговые инфекции имеют короткий инкубационный период, защитными факторами организма при этих инфекциях являются преимущественно секреторные антитела (IgA), а эффективными вакцинами оказываются те, которые применяются местно и стимулируют образование секреторных антител. Примером очаговых инфекций могут быть аденовирусные, парагриппозные, некоторые герпетические инфекции и др.
Очаговая и генерализованная острая инфекция, как явная, манифестная, так и скрытая, инапппарантная, длится относительно непродолжительное время. Но вирус при скрытой острой инфекции также активно репродуцируется и выделяется из организма, человек является источником инфекции, речь идет лишь об особенностях клинического проявления у данного человека. Скрытая инфекция хоть и течет бессимптомно, но оставляет после себя иммунитет.
Персистентная инфекция характеризуется большой продолжительностью взаимодействия вируса и организма (лат. persistentia упорство, постоянство). Персистенция вируса может быть в форме латентной, бессимптомной инфекции с вирогенией за счет интеграции вирусного генома в клеточный, или без нее, при этом вирус не выделяется из организма и клеток. При воздействии ряда активирующих факторов возможен переход латентной инфекции в острую или хроническую. Например, во время вспышки полиомиелита в Румынии медсестра поместила своего ребенка в палату к больным, отделив простынями угол. Ребенок не заболел, чувствовал себя хорошо. Но ему назначили ультрафиолетовое облучение для стимуляции витаминообразования. В результате развились параличи, латентная инфекция перешла в острую. При хронической персистентной инфекции идет медленное выделение вируса (например, герпетическая инфекция, хронические формы вирусных гепатитов и др.). Периоды ремиссии (временное затихание болезни) перемежаются с периодами обострения.
Медленные вирусные инфекции - своеобразная форма взаимодействия некоторых вирусов с организмом, при которой характерен очень длительный инкубационный период (месяцы и годы), медленное, но неуклонное развитие симптомов с летальным исходом. Медленные вирусные инфекции могут вызываться типичными вирусами - коревой подострый склерозирующий панэнцефалит, врожденная краснуха, прогрессирующий подострый краснушный панэнцефалит, ВИЧ-инфекция и др.
Особая форма медленных вирусных инфекций - подострые трансмиссивные губкообразные (спонгиоформные) энцефалопатии. Эти процессы связываются с особыми инфекционными агентами, которые включены в группу так называемых «необычных вирусов». Наиболее изученным среди них является возбудитель скрепи.
Скрепи - очень старая болезнь, легко узнаваемая из точных клинических описаний, сделанных к 18-ому столетию европейскими наблюдателями. Это первый, и более всех изученный представитель группы атипичных медленных инфекций, наблюдающихся у животных и человека. К настоящему времени известны еще семь болезней, имеющих те же диагностические особенности, что и скрепи. Три из этих болезней встречаются у людей, и две из них, болезнь Крейтцфельда-Якоба и синдром Герстманна-Штреусслера - единственные известные передающиеся деменции человека (деменция - приобретенное слабоумие), третья болезнь – куру.
Все изученные заболевания передаются экспериментально путем заражения различных лабораторных млекопитающих. Инкубационный период может быть от 60 дней при самой быстрой экспериментальной модели скрепи, до срока, превышающего естественную продолжительность жизни, которая у мышей и хомяков равна примерно 2 годам. При естественно развивающихся заболеваниях человека продолжительность инкубационного периода может достигать десятилетий. Наследственные факторы хозяина отражаются на продолжительности инкубационного периода (восприимчивости) при одних заболеваниях и не имеют значения при других.
Эти болезни поражают центральную нервную систему (ЦНС), и они неизменно заканчиваются летальным исходом после хронического прогрессирующего течения продолжительностью несколько недель или месяцев. Они характеризуются невоспалительной вакуолизирующей дегенерацией серого вещества ЦНС, поражая нейроны (отсюда и родовое название «губковидные, спонгиоформные энцефалопатии»). Не-воспалительный характер повреждений находится в соответствии с другой характерной особенностью, а именно, что эти инфекции не в состоянии ни вызвать иммунологическую реакцию, ни ослабить иммунологическую реактивность хозяина к другим инфекциям. Это - одна из причин, почему нет никаких лабораторных диагностических тестов на инфекцию с любым из атипичных агентов, и почему вакцинация - не подходящая стратегия для предупреждения или лечения этих болезней в настоящее время. Также не доступны и никакие другие виды лечения.
Новые диагностические критерии для этих болезней были установлены в 1980-ых годах. Экстракты пораженного мозга содержат патологические скрепи-ассоциированные фибриллы (САФ), легко идентифицирующиеся при электронной микроскопии. САФ - амилоидные фибриллы, которые получаются из нормального гликопротеида с молекулярной массой 33-35 кДа, который присутствует во многих неинфицированных тканях. При заболевании скрепи этот белок подвергается едва различимым, но неустановленным посттрансляционным модификациям. В результате он накапливается в мозгу, становится относительно устойчивым к расщеплению протеиназой и приобретает способность образовывать САФ. Меченые антитела легко окрашивают измененный белок (САФ-белок) в срезах мозга, особенно когда он откладывается для формирования ядер внеклеточных амилоидных бляшек.
Гистологически эти амилоидные бляшки походят на те, которые характерны для болезни Альцгеймера, самой частой формы приобретенного слабоумия человека. Однако подчеркивается, что амилоид при болезни Альцгеймера получается из другого белка, чем при скрепи и болезни Крейтцфельд-Якоба. Не имеется также достоверных доказательств, что болезнь Альцгеймера является передающейся.
Лучше всего изучена природа возбудителя скрепи. Заболевание может быть экспериментально пассировано на животных, в том числе на мышах и хомячках. Возбудитель имеет достаточно малые размеры, чтобы проходить через бактериальные фильтры, что позволяет говорить о том, что по размерам он является вирусо-подобным или меньше вирусов. Однако, его инфекционные свойства необычно устойчивы ко многим физико-химическим воздействиям типа высокой температуры и воздействия ионизирующего или ультрафиолетового излучения. Необычная стабильность и иммунологическая нейтральность этих агентов - основание для их описания как «необычных медленных вирусов».
По биологическим свойствам в опытах на мышах легко дифференцируются приблизительно 10 различных штаммов возбудителя скрепи. И в опытах на хомячках, и в опытах на мышах была зарегистрирована мутация, которая, безусловно, не является редким случаем. Поэтому возбудитель скрепи похож на возбудителей других микробных инфекций в проявлении вариации штаммов и мутации. Это означает, что возбудитель скрепи имеет штаммо-специфический геном. Только на основании априорного предположения, геном возбудителя скрепи, вероятно, является нуклеиновой кислотой, хотя это не было доказано.
Характеристика чувствительности возбудителя скрепи к ультрафиолетовому облучению указывает на то, что предполагаемый нуклеиновый геном имеет очень малые размеры. Его оценочный размер в качестве мишени для ионизирующей радиации - меньше 100 000, что может быть недостаточным для кодирования белка, изучаемого с помощью протеаз в качестве необходимого доказательства инфекционности агента скрепи. Это дало основание для выдвижения гипотезы «вирино», которая предлагает, что белок кодируется генами хозяина. Полагают, что «вирино» состоят из небольшого количества нуклеиновой кислоты в комплексе с белком, синтезированным клеткой-хозяином. Недостаточность иммунной реакции к агентам скрепи и ему подобным можно было бы тогда объяснить просто отсутствием чужеродных антигенов.
Таксономически это позволяет поместить «вирино» между истинными вирусами и вироидами.
Вироиды - новый класс субвирусных агентов, характеризующихся кажущимся отсутствием внеклеточной неактивной фазы (вириона) и геномом намного меньшим, чем у известных вирусов. Вироиды как инфекционные агенты - безбелковые, низкомолекулярные односпиральные кольцевые РНК, стойкие к нагреванию и органическим растворителям, но чувствительные к нуклеазам. Вироиды, вначале идентифицированные при болезни картофеля, оказались также причиной некоторых других болезней растений. Возможно, что возбудители некоторых болезней животных и человека, могут оказаться принадлежащими к классу вироидов.
Поскольку единственной молекулой, идентифицированной в препаратах, обладающих инфекционными свойствами, оказался белок, имеется другая гипотеза, что именно сам белок и является инфекционным агентом скрепи. Эта гипотеза - гипотеза «приона».
Прионы (от английских слов “proteinaceous infectious particle”, означающих белковая инфекционная частица) - особые инфекционные агенты, не содержащие нуклеиновой кислоты. Он не теряет своих инфекционных свойств после обработки нуклеазами, но полностью разрушается протеазами. Полагают, что прионы - это особые белки, которые вызывают дерепрессию постоянно существующих в здоровых клетках генов, которые экспрессируются и дают наработку прионов.
Однако трудно объяснить вариабельность штаммов возбудителя скрепи и его мутации на основе представлений о том, что он является белком.
Прионы описываются как инфекционные белки, лишенные нуклеиновой кислоты, в то время как вирино, как считают, состоят из небольшого количества нуклеиновой кислоты, связанной с белком клетки-хозяина. Широко дискутируется вопрос об истинном положении прионов и вирино и отношениях между ними, если таковые вообще имеются.
В настоящее время медленные инфекции привлекают большое внимание, многие вирусные инфекции считаются медленными или могут протекать как медленные инфекции (вирусные гепатиты, СПИД, бешенство, цитомегаловирусные поражения мозга и другие). Предполагается роль медленной вирусной инфекции в заболеваниях человека, этиология которых до настоящего времени не расшифрована : болезнь Паркинсона (вирус гриппа ?), шизофрения (прион ?), атеросклероз (герпесвирус ?) и др.
Кстати, вначале 1996 года мир был встревожен тем, что в Англии возникло заболевание среди коров, при котором наблюдалось губкообразное поражение головного мозга, его в средствах массовой информации назвали " бешенством коров ". Конечно, это заболевание не имеет связи с вирусом бешенства, речь идет о возбудителе прионового типа. Хоть и не было подтвержденных свидетельств о заболеваниях людей при употреблении в пищу говядины от пораженных коров, но с учетом длительности инкубационного периода при медленных инфекциях получить такие данные непросто, поэтому вопрос остается открытым.
В связи с этим инцидентом остро возник вопрос об устойчивости прионов к температурному воздействию. Сообщалось, что при 80 C возбудитель описываемого заболевания коров инактивируется полностью в течение получаса.
Необходимо также остановиться на смешанных вирусных инфекциях. При заражении двумя вирусами одновременно может происходить независимая репродукция обоих вирусов, усиление репродукции одного из вирусов (экзальтация), а также комплементация репродукция одного, дефектного вируса, только в присутствии другого, вируса-помощника. Вирусо-бактериальные инфекции, как правило, протекают более тяжело. Описаны сочетания бруцелл с вирусом омской геморрагической лихорадки, вируса гепатита В с кандидами, энтеровирусов с энтеробактериями, оппортунистические инфекции при СПИДе.