

Литература БФХ / molekuljarnaja biologija kletki v3
.pdf191
Рис. 17-37. Миобласты in vitro пролиферируют, располагаются упорядоченным образом, а затем сливаются, образуя многоядерные мышечные клетки. Микрофотографии живой культуры (в фазовоконтрастном микроскопе) на последовательных стадиях. Фото В сделано при большем увеличении; видна поперечная исчерченность (указана длинной стрелкой), которая появляется, как только начинает развиваться сократительный аппарат. Видны многочисленные ядра в одной клетке (короткие стрелки). (С любезного разрешения Rosalind Zalin.)
перестают делиться, сливаются и дифференцируются. Однако система регуляции сложна. Для осуществления дифференцировки миобласты должны, например, прикрепиться к межклеточному матриксу. При этом процесс слияния является кооперативным: сливающиеся миобласты, видимо, секретируют какие-то неизвестные факторы, побуждающие к слиянию другие миобласты.
17.6.2. Мышечные клетки могут видоизменять свои свойства в результате смены изоформ специфических белков
[35]
Однажды образовавшаяся скелетная мышечная клетка обычно сохраняется до конца жизни животного; при этом она растет, созревает и изменяет свои свойства в соответствии с функциональными потребностями. Геном содержит различные варианты генов, кодирующих многие характерные белки скелетномышечных клеток, причем сплайсинг РНК-транскриптов некоторых генов может происходить по-разному. В результате образуется множество вариантов (так называемых изоформ) белковых компонентов сократительного аппарата. В процессе созревания мышечной клетки подбираются различные сочетания изоформ, приспособленные к меняющимся требованиям в отношении быстроты сокращения и утомляемости у плода, новорожденного и взрослого организма.
В одной и той же мышце можно найти существующие бок о бок мышечные клетки разных типов - каждый тип с особым набором белковых изоформ. У взрослых животных два типа легко распознать даже невооруженным глазом. Красные мышечные волокна, как, например, в темном курином мясе, богаты связывающим кислород белком миоглобином. Белые мышечные волокна, такие как в белом курином мясе, содержат гораздо меньше миоглобина. Различное содержание миоглобина-белка, родственного гемоглобину, - отражает различную потребность в кислороде: для красных волокон более характерно окислительное фосфорилирование, для белых - анаэробный гликолиз. Различные типы метаболизма в свою очередь связаны с разными типами сократительной активности. Красные волокна в ответ на стимуляцию сокращаются медленно, они меньше подвержены утомлению и более эффек-

192
Рис. 17-38. Два последовательных среза одного участка мышцы взрослой курицы, окрашенные флуоресцентными антителами, специфическими для двух разных изоформ миозина. А. Белые (быстро сокращающиеся) клетки окрашены антителами к «быстрому» миозину. Б. Красные (медленно сокращающиеся) клетки окрашены антителами к «медленному» миозину. (G. Gauthieret al., J. Cell Biol, 92, 471-484, 1982.)
тивны при необходимости длительных усилий. Белые волокна дают быстрый ответ, легче утомляются и более эффективны при отдельных быстрых движениях. Такие мышцы, как, например, бицепсы, обычно содержат смесь нескольких типов мышечных клеток в соотношении, оптимальном для функционирования данной мышцы (рис. 17-38).
Изменяя хирургическим путем иннервацию мышц или искусственно стимулируя их вживленными электродами, можно показать, что частот электрического возбуждения сильно влияет на картину экспрессии генов в мышечной клетке. Если «медленную» мышечную клетку стимулировать с частотой, более подходящей для «быстрой» мышцы, то он частично превращается в быструю, и наоборот; это происходит отчасти за счет смены изоформ белков. Как мы уже видели (разд. 17.4.3), такие изменения возможны не только в мышечных клетках: «переключения экспрессии генов, приводящие к появлению вариантов мРНК, чаете происходят при созревании дифференцированной клетки или при ее реакции на окружающие условия.
17.6.3. Некоторые миобласты сохраняются во взрослом организме как покоящиеся стволовые клетки [36]
Мышца может расти тремя способами: возможно увеличение длины дифференцированных мышечных волокон, их толщины и их числа Поскольку клетки скелетной мышцы не способны делиться, новы волокна могут возникать только путем слияния миобластов. Число многоядерных клеток в скелетных мышцах фактически достигает уровня, свойственного взрослому организму, на весьма ранней стадии развития, у человека - еще до рождения. Последующее колоссальное нарастание мышечной массы идет за счет увеличения размеров отдельных клеток. Рост мышцы в длину зависит от включения все большего числа миобластов в существующие многоядерные клетки, главным образом на их концах, что увеличивает число ядер в каждой клетке Напротив, утолщение мышцы, как, например, у штангистов, зависит от увеличения размеров и числа сократительных миофибрилл в каждой отдельной клетке (разд. 11.1.1), а не от изменения числа мышечных клеток или их ядер.
Тем не менее и во взрослом организме сохраняются немногочисленные миобласты. Это маленькие, уплощенные и неактивные клетки, находящиеся в тесном контакте со зрелыми мышечными волокнами и окруженные своей базальной мембраной. В случае повреждения мышцы или при ее обработке фактором роста фибробластов в этих так

193
Рис. 17-39. Радиоавтограф одного многоядерного мышечного волокна с прилегающими клетками-сателлитами. Волокно было выделено у взрослой крысы и перенесено в культуральную среду, содержавшую 3Н-тимидин и экстракт из поврежденной мышцы, который стимулирует деление клеток-сателлитов. Делящиеся сателлиты (показаны стрелками) включили метку (зерна серебра имеют вид черных точек). Ядра мышечных клеток не способны к делению и остаются немеченными. (R. Bischoff, Dev. Biol., 115, 140147, 1986.)
называемых клетках-сателлитах пробуждается пролиферативная активность (рис. 17-39), и их потомки могут сливаться, образуя новые мышечные волокна. Таким образом, эти покоящиеся стволовые клетки в зрелой скелетной мышце находятся в резерве, но при надобности могут служить самообновляющимся источником терминально дифференцированных клеток.
Заключение
Клетки (волокна) скелетных мышц у позвоночных составляют один из четырех видов специализированных клеток, несущих функцию сокращения. Они ответственны за произвольные движения. Каждая клетка представляет собой синцитий и образуется в результате слияния миобластов. Миобласты могут быть стимулированы к пролиферации факторами роста, такими как ФРФ, но после слияния они уже делиться не могут. Слияние миобластов обычно сопряжено с началом дифференцировки мышечной клетки, когда в ней координированным образом включается много различных генов. Впоследствии клетки могут видоизменять свой дифференцированный характер путем изменения набора синтезируемых ими изоформ белков. В мышцах взрослого организма часть миобластов продолжает существовать в состоянии покоя в виде клеток-сателлитов. В случае повреждения мышцы они играют роль стволовых клеток - начинают пролиферировать и сливаться, чтобы возместить утрату мышечных волокон.
17.7. Фибробласты и их превращения: семейство соединительнотканных клеток [37]
Многие дифференцированные клетки взрослого организма можно объединить в семейства, принадлежность к которым определяется происхождением и свойствами клеток. Важный пример - семейство соединительнотканных клеток; члены его не только родственны, но и в необычной степени способны к взаимным превращениям. В это семейство входят фибробласты, хрящевые клетки и костные клетки. Все они специализированы для секреции межклеточного матрикса, содержащего коллаген, и совместно образуют «архитектурный каркас» тела вместе с жировыми клетками и клетками гладкой мускулатуры, имеющими, по-видимому, то же происхождение. На рис. 17-40 изображены эти типы клеток и показаны их возможные превращения. Соединительнотканные клетки играют центральную роль как опорные элементы
Рис. 17-40. Взаимные превращения, по-видимому, происходящие внутри семейства соединительнотканных клеток. Для упрощения фибробласты представлены клеткой одного типа, в действительности же существует много типов фибробластов; число их, однако, не установлено, так же как их потенции к дифференцировке.
194
и участники восстановительных процессов почти в каждой ткани и каждом органе; пластичность их дифференцировки имеет большое значение при реакциях на разного рода повреждения.
17.7.1. Фибробласты изменяют свои свойства в ответ на сигналы от межклеточного матрикса [37, 38]
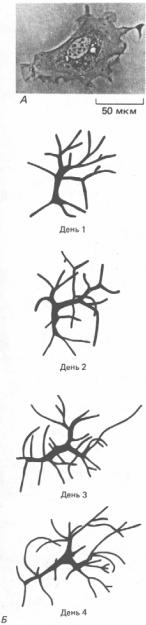
Фибробласты - по-видимому, наименее специализированные клетки в семействе соединительнотканных клеток. В составе соединительной ткани они разбросаны по всему организму и секретируют мягкий внеклеточный матрикс, богатый коллагеном типа I и/или типа III, как это описано в гл. 14 (разд. 14.6.2). В случае повреждения ткани ближние фибробласты мигрируют в рану, размножаются там и образуют большие количества коллагенового матрикса, который помогает изолировать и восстановить поврежденную ткань. Способность этих клеток существовать и действовать в необычных условиях раны наряду с их «одиночным образом жизни» позволяет очень легко выращивать их в культуре, что делает их излюбленным объектом исследований клеточных биологов (рис. 17-41).
Как показано на рис. 17-40, фибробласты - это, видимо, самые пластичные из соединительнотканных клеток: они проявляют удивительную способность дифференцироваться в других представителей того же семейства. Однако, прежде чем углубляться в детали, мы должны предостеречь от слишком поспешных выводов. Есть веские данные в пользу того, что фибробласты в разных частях организма внутренне различны (см. Приложение в конце этой главы); не может быть уверенности даже в том, что в одном участке все фибробласты эквивалентны. Проще всего предположить, что они идентичны, поскольку не доказано, что это не так. Однако вполне возможно, что соединительная ткань содержит смесь различающихся линий фибробластов, из которых одни способны превращаться в хондроциты, другие - в жировые клетки, и т.д.; отсутствие видимых различий не обязательно означает, что существует только один тип фибробластов с множественными потенциями развития. Возможно также, что «зрелые» фибробласты, не способные к трансформации, существуют бок о бок с «незрелыми» (их часто называют мезенхимными клетками), которые могут превращаться в зрелые клетки разного типа.
Несмотря на эти неясности, однако, исследования in vivo и in vitro четко показали, что свойства соединительнотканных клеток могут радикально изменяться. Например, если препарат костного матрикса, полученный размалыванием кости в тонкий порошок и элюированием твердого минерального вещества, имплантировать в дермальный слой кожи, то там некоторые клеточные элементы - возможно, кожные фибробласты - превращаются в хрящевые клетки, а несколько позже другие клетки трансформируются в костные. При этом образуется небольшой кусочек кости, имеющий даже костномозговую полость. Эти результаты позволяют думать, что компоненты внеклеточного матрикса могут сильнейшим образом влиять на дифференцировку соединительно-
Рис. 17-41. А. Фазовоконтрастная микрофотография мышиного фибробласта в культуре. Б. Изображения живой фибробластоподобной клетки в прозрачном хвосте головастика; показаны изменения ее формы и положения в течение нескольких дней. Обратите внимание на то, что, хотя в культуре фибробласты распластываются, в тканях они могут иметь более сложную конфигурацию с многочисленными отростками. (А - с любезного разрешения Guenter Albrecht-Buehler; Б - из Е. Clark, Am.J.Anat., 13, 351-379, 1912.)
195
тканных клеток. Далее мы увидим, что сходные превращения клеток играют важную роль и при срастании костей после переломов. Показано даже, что костный матрикс содержит в себе высокие концентрации нескольких факторов роста, влияющих на поведение соединительнотканных клеток, например TGF-β (трансформирующий фактор роста (3-см. разд. 13.3.4, табл. 13-1), который, как установлено, индуцирует дифференцировку хряща in vitro.
17.7.2. Внеклеточный матрикс может влиять на дифференцировку соединительнотканных клеток, изменяя их форму и способность к прикреплению [39]
Внеклеточный матрикс может влиять на дифференцированное состояние соединительнотканных клеток как физическим, так и химическим путем. Это было показано в исследованиях на хрящевых клетках (хондроцитах), растущих в культуре. При надлежащих условиях эти клетки размножаются и поддерживают свой дифференцированный статус, синтезируя на протяжении многих клеточных поколений большие количества весьма характерного хрящевого матрикса, которым они сами себя окружают. Однако если культуру вести при относительно низкой плотности клеток в виде монослоя на культуральной чашке, то происходит трансформация: клетки теряют округлую форму, типичную для хондроцитов, распластываются на субстрате и перестают вырабатывать хрящевой матрикс. В частности, они прекращают синтез коллагена типа II, характерного для хряща, а вместо этого начинают производить коллаген типа I, характерный для фибробластов. За месяц существования такой культуры почти все хрящевые клетки переключают экспрессию генов коллагена и приобретают вид фибробластов. Биохимические изменения в клетке должны происходить внезапно, так как лишь в очень немногих клетках отмечается одновременный синтез коллагена обоих типов.
Разного рода экспериментальные данные говорят о том, что биохимические изменения по крайней мере частично индуцируются изменением формы клеток и их прикрепления. Например, хондроциты, претерпевшие трансформацию в фибробластоподобные клетки, можно осторожно отделить от поверхности культуральной чашки и перенести на чашку с агарозой. Образуя гель, агароза удерживает клетки в состоянии суспензии, без прикрепления к субстрату, и это заставляет их принимать округлую форму. В таких условиях клетки вскоре вновь приобретают свойства хондроцитов и начинают синтезировать коллаген типа П. Вопрос о том, каким образом форма клеток и их прикрепление могли бы влиять на экспрессию генов, обсуждается в разд. 13.3.6.
Для большинства клеток, особенно соединительнотканных, возможности прикрепления зависят от окружающего матрикса, который обычно вырабатывают сами клетки. Таким образом, клетка создает себе окружение, которое в свою очередь воздействует на клетку, закрепляя ее дифференцированное состояние. Более того, внеклеточный матрикс, образуемый клеткой, отчасти создает окружение и для соседних с ней клеток, способствуя их дифференцировке в том же направлении. Можно наблюдать, например, как в развивающемся организме или в культуральной чашке увеличивается группа хондроцитов, образующая узелок хряща, в результате превращения находящихся рядом фибробластов в хондроциты.

196
Рис. 17-42. Превращение фибробластоподобной клетки-предшественника в зрелую жировую клетку в результате накопления и слияния жировых капелек. Как показано стрелками, этот процесс по крайней мере частично обратим. Клетки, находящиеся в начальной или промежуточной стадии, могут делиться, но зрелая жировая клетка к этому не способна.
17.7.3. Различные сигнальные молекулы, действуя последовательно, регулируют образование жировых клеток [40]
Полагают, что жировые клетки, или адипоциты, у млекопитающих тоже развиваются из фибробластоподобных клеток, как в ходе нормального развития, так и при различных патологиях, например при миодистрофии, когда мышечные клетки погибают и постепенно замещаются жировой соединительной тканью. Дифференцировка жировой клетки начинается с образования специфических ферментов; затем происходит накопление жировых капелек, которые в дальнейшем сливаются и увеличиваются в объеме, настолько сильно растягивая клетку, что остается лишь тонкий ободок цитоплазмы вокруг массы жира (рис. 17-42).
Факторы, влияющие на этот процесс, можно исследовать в культуре клеток, используя клеточные линии, например некоторые штаммы мышиных клеток ЗТЗ. Вначале было обнаружено, что для развития жировых клеток в культуре необходимо присутствие сыворотки плода коровы - обычного компонента культуральной среды. Главный фактор сыворотки, запускающий дифференцировку жировой клетки, позднее был идентифицирован как гормон роста - белок, в норме выделяемый в кровяное русло гипофизом. Получены данные, что этот гормон стимулирует дифференцировку не только жировых клеток, но и хондроцитов, и оказывает такое действие как in vitro, так и in vivo. Но гормон роста - это не единственная секретируемая сигнальная молекула, регулирующая развитие жировых клеток. Предшественники жировых клеток, стимулированные гормоном роста, становятся чувствительными к IGF1 (инсулиноподобный фактор роста 1), который побуждает дифференцирующиеся жировые клетки к пролиферации.
На дифференцировку жировых клеток, так же как и хондроцитов, влияют факторы, изменяющие их форму и прикрепление. Например, дифференцировка клеток ЗТЗ в жировые клетки подавляется, если у них есть возможность распластаться на поверхности культуральной чашки, покрытой фибронектином, к которому они прочно прилипают. Однако это подавление можно снять с помощью цитохалазина, так как он разрушает актиновые нити и в результате клетки округляются.
Все эти эксперименты на клетках соединительной ткани иллюстрируют общее положение: дифференцировку регулирует совместное действие растворимых сигнальных веществ и контактов с внеклеточным матриксом. Эффект каждого фактора зависит от свойств реагирующей клетки, которые в свою очередь зависят от предыстории развития данной клетки.
197
Заключение
В семейство соединительнотканных клеток входят наряду с фибробластами хрящевые, костные, жировые и гладко мышечные клетки. Фибробласты, по-видимому, способны превращаться во все другие клетки этого семейства, и в некоторых случаях это превращение обратимо. Пока не ясно, присуще ли это свойство одному типу фибробластов, которые плюрипотентны, или же речь идет о смеси различных фибробластов с более узкими потенциями. Превращения соединительнотканных клеток регулируются составом окружающего внеклеточного матрикса, формой клеток, гормонами и факторами роста.
17.8. Мягкие клетки и твердый матрикс: рост, обновление и репарация кости [41]
Кость - это очень плотная специализированная соединительная ткань. Подобно железобетону, костный матрикс состоит из двух компонентов - прочных волокон (фибриллы коллагена типа II), противодействующих растяжению, и твердых частиц (фосфат кальция в виде кристалликов гидоксиапатита), устойчивых к сжатию. Объем пространства, занятого коллагеном, почти равен объему, занимаемому фосфатом кальция. В костях взрослого организма фибриллы расположены упорядоченными слоями, напоминая структуру фанеры: в каждом слое они лежат параллельно друг другу, но под прямым углом к фибриллам обоих смежных слоев.
Кость при всей ее твердости подвержена изменениям. Весь ее плотный внеклеточный матрикс пронизан каналами и полостями, заполненными живыми клетками, которые составляют около 15% веса компактной кости. Эти клетки участвуют в непрекращающемся процессе перестройки костной ткани. Клетки одного типа разрушают старый костный матрикс, а клетки другого типа образуют новый. Этот механизм обеспечивает обновление матрикса внутри кости.
Кость может расти только путем аппозиции, т. е. отложения дополнительного матрикса вместе с клетками на свободной поверхности твердой ткани. У эмбриона аппозиционный рост кости должен быть скоординирован с ростом других тканей так, чтобы организм мог увеличиваться в размерах без существенного искажения пропорций. Рост большей части скелета, особенно рост длинных костей конечностей и туловища, координируется с помощью сложной стратегии. У эмбриона сначала из хряща образуются миниатюрные «модели» будущих костей. Каждая такая модель растет, и по мере образования нового хряща более старый хрящ замещается костью. Рост и разрушение хряща и отложение кости в период развития так тонко скоординированы, что у взрослого животного кость, даже достигнув полуметра в длину, имеет почти такую же форму, как первоначальная хрящевая модель, длина которой не превышала нескольких миллиметров. Не углубляясь детально в геометрию этого процесса, мы остановимся на активности клеток, лежащей в основе роста костей и хряща у зародыша и обновления соответствующих тканей у взрослого организма; здесь особенно четко выявляется роль взаимодействий между клетками разного типа.
17.8.1. Хрящ способен к интерстициальному росту [39, 42]
«Сотрудничество» кости и хряща основано на их контрастных свойствах. Обе ткани образуются из мезенхимных клеток, секретирующих большие количества внеклеточного матрикса, который содержит коллаген. Но костный матрикс тверд, а хрящевой может деформироваться, так как

198
Рис. 17-43. А. Схематическое изображение среза, проходящего через хрящевой стержень в период его образования. Показан окружающий хрящевую ткань фиброзный перихондрий. Каждый хондроцит заполняет лакуну в хрящевом матриксе. Б. Поперечный срез через такой стержень у куриного эмбриона на ранней стадии развития. По мере роста ткани количество хрящевого матрикса, приходящегося на один хондроцит, будет сильно возрастать, и граница между хрящом и перихондрием обозначится более резко. (С любезного разрешения Peter Gould.)
состоит в основном из протеогликанов в высокой концентрации (разд. 14.2.4) и из коллагена типа II. Поэтому хрящ в отличие от кости способен к интерстициальному росту: он может увеличиваться в объеме за счет того, что клетки, уже окруженные матриксом, продолжают секретировать его.
Хрящевые клетки, или хондроциты, отделены друг от друга матриксом. Каждая клетка занимает в матриксе небольшую полость, или лакуну. Хрящ обычно не содержит кровеносных капилляров, и жизнедеятельность его клеток поддерживается благодаря диффузии питательных веществ и газов через матрикс от лежащих довольно далеко кровеносных сосудов и в обратном направлении. Большую часть хряща окружает перихондрий - плотный слой соединительной ткани, содержащей коллаген (рис. 17-43). Хрящ растет изнутри, по мере того как хондроциты секретируют новый матрикс, а волокнистый перихондрий выступает в роли корсета, ограничивающего изменения формы. В процессе роста хряща образуются и новые клетки: хондроцит, находящийся в своей лакуне, разделившись, даст две клетки, и каждая из них будет затем секретировать новый матрикс, который вскоре изолирует их друг от друга (рис. 17-44). Новые клетки могут также переходить в хрящ из перихондрия. Как полагают, перихондриальные клетки, напоминающие фибробласты, делятся и подвергаются превращению, в результате которого они начинают образовывать вокруг себя хрящевой матрикс и быстро становятся настоящими хондроцитами. По-видимому, этот процесс, происходящий in vivo, аналогичен описанной выше трансформации соединительнотканных клеток in vitro (разд. 17.7.2).
Рис. 17-44. Рост хряща. Ткань разрастается, по мере того как хондроциты делятся и производят больше матрикса. Новосинтезированный матрикс, непосредственно окружающий клетки, выделен более темным серым цветом. Хрящ может также расти благодаря поступлению фибробластов из перихондрия и их превращению в хондроциты (см. рис. 17-43).

199
17.8.2. Остеобласты секретируют костный матрикс, а остеокласты разрушают его [41, 43]
Кость - ткань более сложная, чем хрящ. Костный матрикс секретируют остеобласты, которые лежат на поверхности уже существующего матрикса и наслаивают на него новый костный материал. Некоторые остеобласты остаются свободными на поверхности, в то время как другие постепенно погружаются в продукт своей собственной секреции. Этот свежеизготовленный материал (состоящий главным образом из коллагена типа I) называется остеоидом. Он быстро превращается в плотный костный матрикс в результате отложения фосфата кальция. Оказавшись заключенной в твердый матрикс, исходная костеобразующая клетка, называемая теперь остеоцитом, уже не имеет возможности делиться, хотя и продолжает секретировать в небольших количествах матрикс. Подобно хондроциту, остеоцит занимает небольшую полость, или лакуну, в матриксе, но в отличие от хондроцитов он не изолирован от своих собратьев: от каждой лакуны отходят очень узкие канальцы, которые содержат отростки лежащего в лакуне остеоцита, позволяющие ему образовывать щелевые контакты с соседними остеоцитами (рис. 17-45). Хотя такие сети из остеоцитов сами не секретируют матрикс и не разрушают его, им, вероятно, принадлежит важная роль в регуляции активности тех клеток, которые это делают.
В то время как остеобласты откладывают костный матрикс, остеокласты разрушают его (рис. 17-46). Остеокласты - это крупные многоядерные клетки типа макрофагов. Подобно другим макрофагам, они развиваются из моноцитов, образующихся в кроветворной ткани костного мозга. Предшественники остеокластов - моноциты - выходят в кровяное русло и скапливаются в местах резорбции кости; там они сливаются друг с другом, образуя многоядерные остеокласты, которые внедряются в поверхностные слои костного матрикса и постепенно растворяют его.
Остеокласты способны проделывать глубокие ходы в материале компактной кости, образуя полости, в которые затем проникают другие клетки. По оси такого туннеля прорастают кровеносные капилляры, а стенки его покрываются слоем остеобластов (рис. 17-47). Остеобласты откладывают концентрическими слоями новую кость, которая постепенно заполняет полость, оставляя лишь узкий канал вокруг нового кровеносного сосуда. Многие остеобласты оказываются замурованными
Рис. 17-45. Эта схема показывает, как остеобласты, выстилающие поверхность кости, секретируют органический матрикс кости (остеоид) и превращаются в остеоциты по мере погружения в этот матрикс. Образующийся матрикс вскоре обызвествляется. Полагают, что сами остеобласты ведут свое происхождение от остеогенных стволовых клеток - близких родственников фибробластов.

200
Рис. 17-46. Остеокласт - гигантская клетка, разрушающая костный матрикс. «Гофрированная каемка»-это место секреции кислот (для растворения минерального вещества кости) и гидролаз (для переваривания органических компонентов матрикса). Клетка показана в поперечном разрезе. Форма остеокластов изменчива, они подвижны и часто посылают отростки для резорбции кости сразу во многих участках. Остеокласты развиваются из моноцитов, их можно рассматривать как специализированные макрофаги. (R. V. Krstic: Ultrastucture of the Mammalian Cells An Atlas. Berlin: Springer, 1979.)
в костный матрикс и образуют там концентрические кольца остеоцитов. В то время как одни туннели заполняются костью, другие заново прокладываются остеокластами в более старых концентрических системах. Результаты этой непрерывной перестройки хорошо видны на гистологических препаратах компактной кости (рис. 17-48).
Рис. 17-47. Схема, иллюстрирующая процесс перестройки компактного вещества кости. Остеокласты, действуя небольшими группами, прокладывают тоннели в старой кости, продвигаясь ежедневно приблизительно на 50 мкм. Вслед за ними в тоннель входят остеобласты; они выстилают стенки тоннеля и начинают образовывать новую кость, откладывая матрикс со скоростью 1-2 мкм в сутки. Одновременно по оси тоннеля прорастают капилляры. В конце концов тоннель будет постепенно заполняться концентрическими слоями новой кости - останется свободным только узкий центральный канал. Каждый такой канал не только обеспечивает доступ остеобластам и остеокластам, но содержит также один или несколько кровеносных сосудов, доставляющих питательные вещества, необходимые для жизни костных клеток. Обычно за год у здорового взрослого млекопитающего таким способом заменяется 5-10% кости. (По Z. F. G. Jaworski, В. Duck, G. Sekaly, J. Anat., 133, 397-405, 1981.)