

Литература БФХ / molekuljarnaja biologija kletki v3
.pdf1
2

3
Б. Албертс Д. Брей Дж. Льюис М. Рэфф К. Робертс Дж. Уотсон
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ КЛЕТКИ
2-Е ИЗДАНИЕ, ПЕРЕРАБОТАННОЕ И ДОПОЛНЕННОЕ
В 3 томах
3
Перевод с английского канд. биол. наук В. П. Коржа,
канд. биол. наук Н.В. Сониной, канд. биол. наук Н. М. Руткевич, канд. биол. наук Г. И. Эйснер,
Т. Д. Аржановой, Г. В. Крюковой, А. В. Никашина
под редакцией акад. Г. П. Георгиева, д-ра биол. наук Ю.С. Ченцова
Москва |
1994 |
4
ББК 28.070 М75 УДК 576.32/36
Издание выпущено в счет дотации, выделенной Комитетом РФ по печати
Авторы: Албертс Б., Брей Д., Льюис Дж., Рэфф М., Роберте К., Уотсон Дж.
Молекулярная биология клетки: В 3-х т. 2-е изд., М75 перераб. и доп. Т. 3. Пер. с англ.-М.: Мир, 1994.-504 с., ил. ISBN 5-03-001985-5
Созданный коллективом известных американских ученых (в их числе - лауреат Нобелевской премии Джеймс Уотсон) современный учебник молекулярной биологии. Энциклопедическая полнота охвата материала позволяет использовать его как справочное пособие. На русском языке выходит в 3-х томах. Читатель уже знаком с 1-м изданием (М.: Мир, 1986-1987). Новое издание переработано авторами и дополнено современным материалом. В т. 3 рассматриваются проблемы клеточной дифференцировки и организации специализированных тканей, анализируются общебиологические и молекулярно-генетические аспекты злокачественного перерождения клеток.
Для биологов всех специальностей, преподавателей и студентов университетов, медицинских, педагогических и сельскохозяйственных институтов.
Учебное издание
Брюс Албертс, Деннис Брей, Джулиан Льюис, Мартин Рэфф, Кейт Робертс Джеймс Д. Уотсон
МОЛЕКУЛЯРНАЯ
БИОЛОГИЯ
КЛЕТКИ
2-е издание, переработанное и дополненное
В 3-х томах
Том 3
ISBN 5-03-001988-Х (русск.)
ISBN 5-03-001985-5
ISBN 0-8240-3965-6 (англ.)
ББК 28.070
Лицензия Л.Р. № 010174 от 22.01.92 г.
Зав. редакцией канд. биол. наук М. Д. Гроздова Ведущие редакторы: Ю. И. Лашкевич, М. Б. Николаева, М. Р. Погосбекова Редактор Н. В. Пересыпкина Художник Е. И. Волков
Художественные редакторы А. Е. Волков, Л. М. Аленичева Технический редактор М. А. Страшнова Корректор Н. Н. Светлова
ИБ № 7581
Сдано в набор 19.11.92г. Подписано к печати 21.09.94г. Формат 84 х 108 1/16. Бумага офсетная. Печать офсетная. Гарнитура таймс. Объем 15,75 бум. л. Усл. печ.л. 52,93.
Усл.кр.-отт. 103,74. Уч.изд.л. 55,25. Изд. № 4/7793. Тираж 10000 экз. Зак. 2292.
Издательство «Мир» Комитета Российской Федерации по печати 129820, ГСП, Москва, И- 110, 1-й Рижский пер., д. 2
Можайский полиграфкомбинат Комитета Российской Федерации по печати 143200, г. Можайск, ул. Мира, 93
Редакция литературы по биологии
©1989 by Bruce Alberts, Dennis Bray, Julian Lewis, Martin Raff, Keith Roberts, and James D. Watson
©перевод на русский язык, Корж В. П., Сонина Н. В., Руткевич Н.М., Эйснер Г. И., Никашин А. В., Аржанова Т. Д., Крюкова Г. В.
1994

5
III От клеток к многоклеточным организмам
15-дневный эмбрион мыши
6
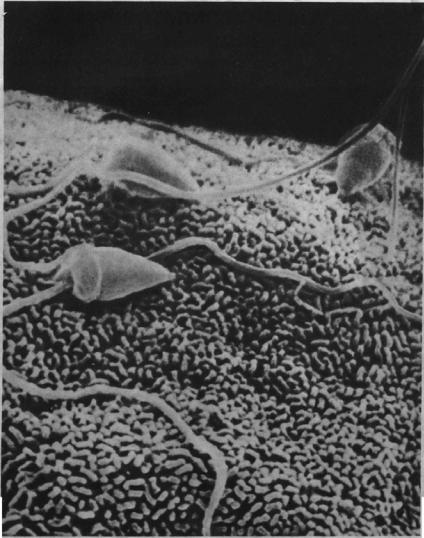
Сперматозоид на поверхности яйцеклетки морского ежа. Фотография, полученная с помощью растрового электронного микроскопа. (С любезного разрешения Brian Dale.)

7
15 Половые клетки и оплодотворение
Размножение возможно и без полового процесса. Одноклеточные организмы могут размножаться простым митотическим делением. Многие растения размножаются вегетативным путем, образуя многоклеточные дочерние особи, которые впоследствии отделяются от материнского организма. Подобные явления имеют место и в животном царстве: так, одиночная многоклеточная гидра производит потомков, отпочковывая их от своего тела (рис. 15-1). Актинии и некоторые морские черви делятся на две половинки, каждая из которых регенерирует недостающую часть организма. У ящериц встречаются даже такие виды, которые представлены лишь женскими особями и размножаются без спаривания. Такого рода бесполое размножение - процесс весьма несложный и не ведет к образованию новых форм: все потомство генетически идентично родительскому организму. В отличие от этого при половом размножении происходит смешивание геномов двух разных особей данного вида, и образующиеся в результате потомки обычно генетически отличаются друг от друга и от обоих родителей. Половое размножение, по-видимому, имеет большие преимущества, так как оно свойственно подавляющему большинству растений и животных. Даже многие прокариоты и некоторые другие организмы, обычно размножающиеся неполовым путем, в некоторых случаях переходят к половому размножению, в результате которого возникают новые комбинации генов. В этой главе мы познакомимся с клеточными механизмами, функционирующими при половом размножении, но прежде чем переходить к подробностям, мы рассмотрим причины возникновения этого аппарата и преимущества, которые он дает.
15.1. Преимущества полового процесса
Цикл полового размножения включает чередование гаплоидных поколений клеток, каждая из которых имеет одинарный набор хромосом, с диплоидными поколениями, где клетки обладают двойным хромосомным набором (рис. 15-2). Смешивание геномов происходит благодаря слиянию двух гаплоидных клеток, из которых образуется одна диплоидная. В свою очередь новые гаплоидные клетки образуются из диплоидных в результате деления особого типа, называемого мейозом. В процессе генетической рекомбинации в мейозе парные хромосомы обмениваются ДНК, после чего новые их комбинации расходятся в разные клетки, которые теперь содержат одинарные наборы хромосом (см. разд. 15.2.2). В результате каждая клетка нового гаплоидного поколения получает новое сочетание генов, происходящих частично от одной родительской клетки предыдущего гаплоидного поколения и частично от другой. Таким образом, благодаря циклам, включающим гаплоидную фазу, слияние гамет, диплоидную фазу и мейоз, распадаются старые комбинации генов и создаются новые.
Рис. 15-1. Гидра, от которой отпочковываются две новые особи (указаны стрелками). Потомки генетически идентичны родительскому организму; они в конце концов, отделяются и переходят к независимому существованию. (С любезного разрешения Matai Hornbruch.)

8
15.1.1.У многоклеточных животных диплоидная фаза бывает сложной и продолжительной, а гаплоидная - простой
икратковременной
В ходе полового цикла клетки размножаются путем обычного митотического деления - чаще всего во время диплоидной фазы (см. разд. 13.5). Исключение составляют некоторые простые организмы, например дрожжи (путем митоза у них размножаются только гаплоидные клетки, диплоидная же клетка, образовавшись, сразу переходит к мейозу), а также растения, хотя и не в столь яркой форме; у последних митотические деления происходят и в гаплоидной, и в диплоидной фазах. При этом у всех растений, за исключением наиболее примитивных, гаплоидная фаза очень короткая и простая, тогда как диплоидная представлена длительным периодом развития и роста. Почти у всех многоклеточных животных, и в том числе у всех позвоночных, гаплоидная фаза еще короче. Практически весь свой жизненный цикл они проводят в диплоидном состоянии; гаплоидные клетки живут очень недолго, они совсем не делятся и специально приспособлены для полового слияния (рис. 15-3).
Гаплоидные клетки, которые сливаются при оплодотворении, называются гаметами. В типичном случае образуются гаметы двух типов: крупные неподвижные яйцеклетки (или яйца) и мелкие, способные передвигаться спермии (или сперматозоиды) (рис. 15-4). Во время диплоидной фазы, начинающейся сразу после слияния гамет, клетки размножаются и специализируются, образуя сложный многоклеточный организм. У большинства животных (но не растений) полезно различать клетки зародышевой линии (зачаткового пути), от которых берет начало
Рис. 15-2. Жизненный цикл организма, размножающегося половым путем, включает чередование диплоидного поколения клеток с гаплоидным.
Рис. 15-3. Эта схема показывает, как размножаются в диплоидной фазе клетки высших эукариот, образуя многоклеточный организм, в котором гаплоидными становятся только гаметы. Напротив, у некоторых низших эукариот размножаются именно гаплоидные клетки, а единственной диплоидной клеткой является зигота, которая существует очень недолго после оплодотворения. Гаплоидные клетки выделены цветом
9
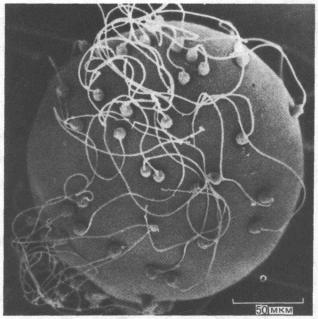
Рис. 15-4. Яйцеклетка двустворчатого моллюска с многочисленными спермиями, прикрепившимися к ее поверхности. Микрофотография получена с помощью сканирующего электронного микроскопа. (С любезного разрешения David Epel.)
следующее поколение гамет, и соматические клетки, образующие весь остальной организм и не оставляющие потомства. В некотором смысле соматические клетки нужны только для того, чтобы способствовать выживанию и размножению клеток зачаткового пути (половых клеток).
15.1.2. Половое размножение делает организмы конкурентоспособными в условиях изменчивости окружающей
среды
Аппарат полового размножения сложен, и средства, «затрачиваемые» на него, очень велики. Какие же преимущества он дает, и почему он выработался в процессе эволюции? При наличии генетической рекомбинации родительские особи производят потомков, которые будут отличаться от них самым непредсказуемым образом, причем среди новых случайных сочетаний генов, по крайней мере, половина может оказаться хуже родительских генотипов. Но если это так, то почему половое размножение должно быть выгоднее бесполого, при котором потомки будут сохранять все родительские гены? Хотя для специалистов по генетике популяций этот вопрос все еще до конца не ясен, основной вывод, повидимому, состоит в том, что перетасовка генов в процессе полового размножения способствует выживанию вида при изменении условий среды. Если родительская особь производит много потомков с самыми разнообразными комбинациями генов, имеется больше шансов на то, что хотя бы один потомок окажется хорошо приспособленным для будущих жизненных обстоятельств, какими бы они ни были. Для объяснения преимуществ полового размножения в борьбе за существование было предложено много гипотез. Одна из них дает представление о том, какими могли быть первые этапы эволюции полового размножения. Ход эволюции в значительной мере зависит от мутаций, которые изменяют существующие гены, образуя вместо них новые аллели (варианты) этих генов. Предположим, что у двух особей в некоторой популяции возникли благоприятные мутации, затрагивающие разные генетические локусы, а значит, и разные функции.
10
У бесполого вида каждая из этих особей даст начало клону мутантных потомков, и два новых клона будут конкурировать до тех пор, пока один из них не одержит верх. Один из благоприятных аллелей, появившихся благодаря мутациям, будет, таким образом, распространяться, тогда как другой в конце концов исчезнет. Теперь представим себе, что один из исходных мутантов обладает генетически обусловленной особенностью, позволяющей ему время от времени включать в свой геном гены из других клеток. В условиях борьбы за существование приобретение генов у клеток конкурирующего клона равносильно созданию клетки, несущей все благоприятные мутации. Такая клетка будет обладать наибольшей приспособленностью, и полученные ею преимущества обеспечат распространение в популяции особенности, позволяющей включать в свой геном гены других клеток. Естественный отбор будет благоприятствовать такому примитивному половому размножению.
Какими бы ни были истоки полового размножения, поразительно то, что эволюция практически всех сложных организмов, доживших до наших дней, протекала благодаря чередованию многих поколений, размножавшихся половым путем. Несмотря на обилие бесполых организмов, они, по-видимому, остались весьма примитивными. Почему? Возможно, ответ состоит в том, что половое размножение создает особые возможности для генетического обновления, ведущего к развитию сложных организмов. Обо всем этом мы будем говорить дальше.
15.1.3. Новые гены появляются в результате дупликаций и дивергенции
Для эволюции сложного организма требуется нечто большее, чем улучшение уже имеющихся генов: нужны новые гены для осуществления новых функций. Как они появляются?
Многие белки многоклеточного животного могут быть сгруппированы в семейства: коллагены, глобины, сериновые протеазы и т. п. Белки одного семейства близки как по своей функции, так и по аминокислотной последовательности. Вряд ли можно сомневаться в том, что гены белков каждого такого семейства произошли от единственного предкового гена в результате процессов дупликации и дивергенции (разд. 10.5.3). Разные члены одного семейства белков часто бывают характерны для различных тканей тела, где они выполняют похожие, но несколько различающиеся функции. Создание новых генов благодаря дивергенции и специализации имеющихся играло, очевидно, решающую роль в эволюции сложных многоклеточных организмов. В этом отношении диплоидные организмы обладают важным преимуществом: у них имеется добавочная копия каждого гена, и эта копия может мутировать и служить исходным материалом для создания чего-то нового. Гаплоидные виды не могут так же легко вступать на путь, ведущий к увеличению и усложнению генома. Чтобы механизм этих процессов стал ясен, нам нужно будет несколько подробнее рассмотреть связь между половым размножением и диплоидией.
15.1.4. Половое размножение сохраняет диплоидность у диплоидных видов
У диплоидного организма имеются две копии каждого гена - по одной от каждого из родителей; однако для выживания и нормальной жизнедеятельности в большинстве случаев бывает достаточно одной копии. Мутация, нарушающая функцию жизненно важного гена, для гаплоидного организма летальна, но она может оказаться безвредной