

Лекция 1.5. Топография активных центров простых и сложных ферментов
Любая ферментативная реакция начинается с взаимодействия субстрата, в большинстве случаев, небольшой по размерам молекулы, с активным центром фермента. Под активным центром фермента понимают совокупность аминокислотных остатков, осуществляющих связывание (сорбцию) субстрата, его химическую активацию и превращение. Активный центр белковой молекулы фермента имеет сложную конфигурацию; он включает как полярные (гидрофильные), так и неполярные (гидрофобные) группы.
Структура активного центра фермента складывается из двух составляющих:
1) сорбционного участка (подцентра, сайта), ответственного за связывание, фиксацию и ориентацию субстратов; свойства этого центра определяют специфичность действия фермента;
2) каталитического участка (подцентра, сайта), осуществляющего химическое превращение молекул субстрата и использующего для этих целей, как правило, общий кислотно-основной катализ.
Аминокислотные остатки, образующие каталитический центр однокомпонентного фермента, расположены в различных точках единой полипептидной цепи. Поэтому активный центр, представляющий собой уникальное сочетание нескольких аминокислотных остатков, возникает в тот момент, когда белковая молекула приобретает присущую ей третичную структуру. Чаще всего в активных центрах однокомпонентных ферментов встречаются остатки Ser, His, три, Arg, Cys, Asp, Glu и Tyr. Изменение третичной структуры фермента под влиянием тех или иных факторов может привести к деформации активного центра и изменению ферментативной активности.
Активный центр двухкомпонентных ферментов представлен небелковым компонентом – коферментом (простетической группой) и несколькими выше приведенными минокислотными остатками.
Характерной особенностью cложных или двухкомпонентных ферментов является то, что ни белковая часть, ни добавочная группа в отдельности не обладают заметной каталитической активностью. Только их комплекс проявляет ферментативные свойства. При этом белок резко повышает каталитическую активность добавочной группы, присущую ей в свободном состоянии в очень малой степени; добавочная же группа стабилизирует белковую часть и делает ее менее уязвимой к денатурирующим агентам. Таким образом, хотя непосредственным исполнителем каталитической функции является простетическая группа, образующая каталитический центр, ее действие немыслимо без участия полипептидных фрагментов белковой части фермента.
В апоферменте есть участок, характеризующийся специфической структурой, избирательно связывающий кофермент. Это так называемый кофермент связывающий домен; его структура у различных апоферментов, соединяющихся с одним и тем же коферментом, очень сходна. Таковы, например, пространственные структуры нуклеотидсвязывающих доменов ряда дегидрогеназ (рис. 1.5.1).

Рис. 1.5.1. Активный центр глюкозо-6-фосфатдегидрогеназы
Методы изучения активных центров ферментов
Представление об активном центре сформировалось в результате анализа данных по ингибированию реакций и химической модификации белковой молекулы. Необратимые ингибиторы блокируют каталитическую активность фермента, осуществляя химическую модификацию одной из групп, участвующих в каталитическом превращении субстрата. Обратимые ингибиторы, образуя комплекс с функциональной группой белка, вызывают либо существенное изменение свойств данной группы (неконкурентные ингибиторы), либо конкурентно блокируют сорбцию (комплексообразование) субстрата в области каталитического центра.
Рассмотрим некоторые примеры.
Сериновые протеазы и эстеразы. Каталитически активной группой многих ферментов является гидроксильная группа серина. В активном центре эта спиртовая группа играет роль нуклеофильного реагента в реакциях нуклеофильного замещения при гидролизе сложных эфиров, амидов, пептидов. Представителем семейства сериновых протеаз является простагландин-Н-синтаза, участвующая в метаболизме арахидоновой кислоты.
Простагландин-Н-синтаза. Аспирин (ацетилсалициловая кислота) представляет собой нестероидный противовоспалительный лекарственный препарат. Физиологическое действие препарата связано с его способностью ацетилировать Ser-514, входящий в центр сорбции арахидоновой кислоты - субстрата ПНС.

Рис. 1.5.2. Блокирование гидроксильной группы серина в активном центре простагландин-Н-синтазы
Аспирин выступает необратимым ингибитором лимитирующего фермента синтеза простагландинов. Последующий гидролиз модифицированного белка и анализ продуктов гидролиза позволили идентифицировать центр модификации фермента.
Несмотря на то, что метод химической модификации позволяет получить весьма важную информацию о природе активных центров ферментов, он имеет и определенные недостатки.
Функциональные группы белка, составляющие активный центр, могут быть замаскированы полипептидной цепью или остатками других аминокислот, что делает группы активного центра недоступными для реагента-модификатора. Химическая модификация, как правило, не является избирательной, химической реакции подвергаются сразу несколько аминокислотных остатков в белке. Это ведет к существенному изменению структуры белка, развитию инактивационных и денатурационных процессов, что может привести к потере ферментом каталитической активности даже в том случае, если химически модифицировались остатки, не входящие в каталитический центр. Выводы об участии тех или иных функциональных групп аминокислот в каталитическом процессе на основе данных по химической модификации белка могут быть сделаны с известной осторожностью и оговорками.
Таким образом, метод химической модификации не позволяет получить исчерпывающую информацию об участниках каталитического акта.
Как правило, для такого рода выводов требуются независимые структурные исследования.
Ситуация становится более однозначной, если химический модификатор встраивается в структуру специфического субстрата или ингибитора фермента. В этом случае модификатор адресно направляется в активный центр, что существенно увеличивает вероятность химической реакции с функциональной группой активного центра.
Новые возможности идентификации групп, входящих в активные центры ферментов, появились с развитием техники сайт-специфического мутагенеза. Для ферментов, экспрессию генов которых можно организовать с помощью генно-инженерных конструкций типа плазмид, оказалась возможной замена отдельных аминокислот на уровне ДНК с последующей экспрессией и изучением каталитических свойств получаемых белков. Это позволяет получить важную информацию об участии той или иной аминокислоты данного фрагмента полипептидной цепи в каталитическом акте. Однако и в этом случае при интерпретации результатов необходима известная осторожность, поскольку в белках имеется большое число аминокислот, формирующих структуру активного центра, но не принимающих непосредственное участие в акте катализа.
Окончательная информация о структуре активного центра активного центра может быть получена методом рентгеноструктурного анализа (РСА) и спектроскопии ядерного магнитного резонанса (ЯМР) высокого разрешения. В первом случае исследование проводят на кристаллах фермента, во втором ‒ исследуют растворы фермента. Для идентификации групп, принимающих участие в катализе, обычно используют образование комплекса ферментов с ингибиторами или мало реакционноспособными аналогами субстратов (т.н. квазисубстратами).
Метод РСА впервые был использован Липскомбом с сотрудниками при анализе активного центра карбоксипептидазы А. На рис. 1.5.3. показана структура карбоангидразы по данным рентгеноструктурного анализа.

Рис. 1.5.3. Третичная структура карбоангидразы по данным рентгеноструктурного анализа: а) общий вид ферментной глобулы; б) пространственное расположение аминокислотных остатков
Структуру и свойства каждого белка определяет последовательность аминокислот. В настоящее время становится очевидным, что при большой вариабельности белков некоторые элементы структуры являются консервативными, и эти элементы в значительной степени определяют функцию белковой молекулы. Это особенно характерно для белков, выполняющих каталитическую функцию. Например, для гидролаз, составляющих около трети всех известных ферментов (приблизительно 1100 из 3700), типов структур каталитических центров всего четыре.
Чтобы ответить на вопросы, какие химические структуры образуют каталитический центр, каким образом аминокислоты, расположенные на разных, зачастую удаленных друг от друга участках полипептидной цепи, находят друг друга и формируют уникальную структуру, ‒ используют методы биоинформатики.
По мнению энзимологов в рамках одного суперсемейства ферментов сорбционный сайт, отвечающий за специфичность, может быть представлен многими вариантами аминокислотных остатков, соответствующими вариантам структуры субстратов. В то же время каталитические сайты, число типов которых весьма ограничено, являются консервативными (незаменимыми) элементами структуры. Для подтверждения этого положения был использован биоинформационный подход, основанный на сравнении последовательностей аминокислот в белках, объединенных в одно крупное семейство.
Был проведен анализ нескольких больших семейств ферментов, представленных в базе данных HSSP (www.sander.embl-heidelberg.de/). Выбор семейств ферментов был сделан на основании следующих критериев:
1) число анализируемых представителей семейства должно быть более 100; это необходимо для обеспечения статистической достоверности результатов;
2) для анализа следует выбирать семейства ферментов различных классов (оксидоредуктазы, гидролазы, изомеразы и т.д.);
3) по возможности следует выбирать ферменты, для которых установлена структура активных центров и с высокой степенью достоверности изучен механизм катализа.
Проведенный анализ показал, что в полипептидной цепи большая часть позиций аминокислот высоко вариабельна, это означает, что функционирование фермента не зависит от того, какую позицию занимает та или иная аминокислота. В то же время имеются позиции аминокислот, которых относительно немного. Эти позиции и соответствующие им аминокислоты называют консервативными. Именно они играют особую роль в функционировании фермента. Что же это за аминокислоты, и какова их роль?
Биоинформационный анализ ферментов всех классов показал, что наиболее часто консервативной аминокислотой является глицин. По рейтингу консервативности аминокислоты располагаются в следующем ряду: глицин > аспарагиновая кислота > цистеин > пролин > гистидин > аргинин > глутаминовая кислота. Это наиболее важные аминокислоты в ферментативном катализе. В сумме глицин и аспарагиновая кислота составляют примерно 50% всех консервативных аминокислот. Из наиболее часто встречающихся консервативных элементов структуры ферментов можно отметить глицин, аспарагиновую кислоту, цистеин, пролин и гистидин. Эти аминокислоты составляют примерно 70% всех консервативных элементов. Метионин и изолейцин практически никогда не бывают консервативными.
В свою очередь наиболее консервативные аминокислоты можно разделить на две принципиально разные группы:
1) аминокислоты, участвующие в активации молекул субстрата в качестве кислот и оснований (аспарагиновая кислота и гистидин);
2) аминокислоты, формирующие геометрию активного центра (глицин, цистеин, пролин).
Таким образом, статистический анализ показал, что каталитическую функцию фермента и архитектуру активного центра формирует небольшая, но определенная часть аминокислот, занимающих строго фиксированные позиции в полипептидной цепи. Консервативные аминокислоты являются либо кислотами или основаниями (электрофильные и нуклеофильные агенты), формирующими каталитический сайт, либо важными структурообразующими аминокислотами, формирующими структуру белка в целом.
Каталитическую функцию выполняют аспарагиновая кислота, гистидин, аргинин, и глутаминовая кислота. Структурообразующими аминокислотами являются глицин, цистеин, и пролин. Глицин и пролин, обеспечивающие возможность поворота цепи, необходимы для того, чтобы активный центр был образован аминокислотами, расположенными на разных участках полипептидной цепи. А цистеин необходим для фиксации необходимой конформации полипептидной цепи.
Природа сформировала активные центры ферментов из ограниченного числа компонентов. Большая часть активных центров ферментов всех классов сформирована из аспарагиновой и глутаминовой кислот, из гистидина и аргинина, из ионов нескольких металлов. Как следствие, число типов каталитических центров невелико. Например, для гидролаз, составляющих около трети всех известных ферментов, можно идентифицировать всего четыре основных типа структуры. Эффективные комбинации каталитических групп, характерные для одних реакций, природа активно использует для организации каталитических центров других типов реакций.
Полипептидная цепь обеспечивает организацию каталитических групп в активные центры. Как известно, в растворе практически исключены трехмолекулярные реакции и реакции более высоких порядков. В ферментативных процессах в реакции участвуют четыре (или пять) остатков различных аминокислот, организованных в полипептидную цепь. Ферментативный катализ не использует сильных химических агентов; компоненты, составляющие активные центры, ‒ это относительно слабые кислоты и основания. Однако они хорошо организованы в пространстве и, как следствие, весьма эффективны.
Примеры активных центров некоторых ферментов
Остановимся на ферментах класса гидролаз, для большинства которых идентифицированы группы, составляющие каталитически активные центры, и созданы обоснованные представления о взаимодействии этих групп в механизме каталитического цикла.
По структуре активных центров и механизму действия гидролазы условно можно разделить на 4 основных типа.
1. Гидролазы, содержащие в активном центре аспарагиновую или глутаминовую кислоту (лизоцим-пепсиновый тип).
2. Гидролазы, содержащие в активном центре гидроксильную группу серина, треонина или цистеина и цепь переноса протонов, активирующую эту группу (тип химотрипсина); гидролазы, использующие имидазольную группу гистидина непосредственно для активации воды (тип панкреатической рибонуклеазы).
3. Гидролазы, использующие комплексы Zn2+ или Со2+ для активации воды и субстрата (тип щелочной фосфатазы, карбоксипептидазы А).
4. Гидролазы, использующие ионы Мg2+ или Мn2+ для активации воды и субстрата (тип пирофосфатазы).
Химотрипсин. В активный центр входят Ser-195, His-57, Asp-102.

Рис. 1.5.4. Структура химотрипсина
Лактатдегидрогеназа. Это NAD+-зависимая дегидрогеназа. Осуществляет обратимое окисление-восстановление органических молекул, при этом в качестве донора (акцептора) гидрид-иона выступает кофермент. Каталитически активные группы фермента представлены Arg-165, His-194, Arg-105. Все эти аминокислоты являются консервативными. Молочная или пировиноградная кислоты фиксируются в активном центре с помощью положительного заряда Arg-168. Участниками каталитического процесса являются протон-транспортная цепь His-194-Asp-165 и Arg-105.
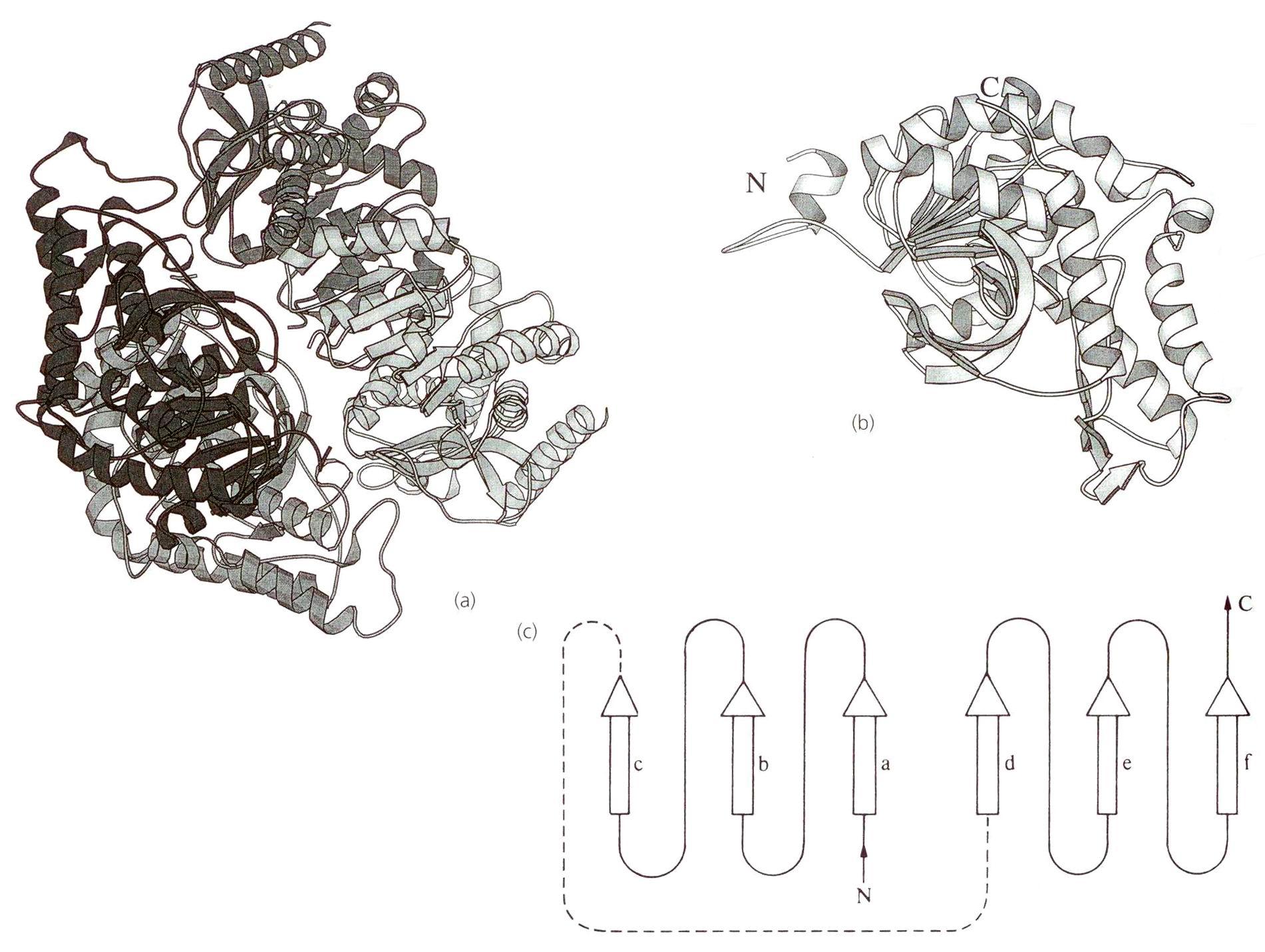
Рис. 1.5.5. Структура лактатдегидрогеназы
(а) Схематическое изображение тетрамера и (b) - отдельной субъединицы; (с) Модель NAD+-связывающего региона. Никотинамидное кольцо NAD+связывается между цепями d и е, а адениновое кольцо – между а и b.
На рис. 1.5.6. приведены возможные типы связей, участвующих в присоединении NAD+ в активном центре ЛДГ.

Рис. 1.5.6. Связывание NAD+ лактатдегидрогеназой
Линии, показанные точками – водородные связи, перекрестные линии – электростатические взаимодействия, аминокислотные остатки в рамках – гидрофобные взаимодействия
Триозофосфатизомераза. Каталитически важные группировки активного центра фермента представлены Glu-165 и His-95.

Рис. 1.5.7. Структура субъединицы триозофосфатизомеразы дрожжей
Глицин, цистеин и пролин как структурообразующие аминокислоты
Глицин в силу особенностей его строения не участвует в химических актах активации молекул в каталитическом цикле. Не обладая заместителем у α-углеродного атома, глицин лишен выраженной химической функции. Тем не менее наличие глицина в структуре белка очень важно. Так, сайт-специфическая замена глицина в консервативных позициях на любую из аминокислот приводит, как правило, к полной потере (или существенному снижению) активности фермента.
По-видимому, глицин в консервативных позициях важен по следующим причинам.
1. Являясь уникальной аминокислотой с наиболее энергетически облегченным вращением вокруг связей С-N и С-С полипептидной цепи, глицин может играть роль узловой точки, обеспечивающей возможность изменения направления полипептидной цепи при «сборке» аминокислотных остатков в активный центр. Таким образом, наличие консервативных глицинов позволяет объяснить структурный парадокс ферментативного катализа, когда одинаковые активные центры «собираются» из абсолютно разных полипептидных цепей. Общим для этих цепей являются наличие глицина в консервативных позициях и возможность стабилизации собранной структуры, например, за счет дисульфидных связей (цистеин также проявляет высокую степень консервативности, занимая третью позицию в рейтинге консервативности).
2. Глицин в консервативных позициях может играть роль конформационных «шарниров», обеспечивая возможность «сборки» активного центра и известную конформационную подвижность. Подтверждением этому служит то, что во многих случаях вблизи каталитически активных групп можно обнаружить глицин в консервативных позициях. Например, для гидролаз различных семейств консервативными являются следующие мотивы: Asp-215-X-Gly-217 (пепсин); Asp-170-Xаа-Xаа-Gly-173 (термолизнн); Gly-173-Xаа-Ser-177 (трипсин); His-76-Gly-77, Ser-153-Xаа-Gly-155, Gly-175-Xаа-Asp-177 (липазы). Здесь Хаа ‒ произвольная аминокислота. Аминокислоты Asp, His, Ser в указанных ферментах входят в структуру активных центров.
Превращение исходного субстрата в конечные продукты в ферментативном катализе сопряжено с участием большого числа интермедиатов с отличной от исходного субстрата структурой. Глицины активного центра могут играть роль «релаксирующих» элементов, конформационно подстраивая активный центр для следующего элементарного акта.
Существенную роль в формировании архитектуры активного центра играют цистеин и пролин (соответственно 3-я и 4-я позиции в рейтинге консервативных аминокислот). Пролин, как известно, является уникальной аминокислотой, разворачивающей полипептидную цепь. Роль цистеина заключается в том, что необходимая конформация активного центра, складывающаяся из различных участков полипептидной цепи, фиксируется химической связью в виде дисульфидного мостика. Для многих ферментов это завершает формирование архитектуры активного центра.
Таким образом, активный центр состоит из ряда функциональных групп, определенным образом ориентированных в пространстве. Среди них различают группы, входящие в состав каталитического сайта активного центра, и группы, образующие сайт, обеспечивающий специфическое сродство, т.е. связывание субстрата ферментом – так называемый контактный или «якорный» участок. Это деление достаточно условно, поскольку взаимодействия в контактном участке фермента при образовании фермент-субстратного комплекса, оказывает существенное влияние на скорость и направление превращений в каталитическом участке.