

3.2. Дослідження ауксотрофності штамів-продуцентів лізину
Згідно методики представленої в роботі [158] встановлення ауксотрофності проводили на твердих поживних середовищах.
Позитивний контроль дослідів отримували на повноцінному середовищі МПАзб., яке містило всі необхідні ростові речовини (рис. 3.3 а). В негативному контролі — МС – ростові речовини були відсутні. Клони, які росли при позитивному контролі, не росли на МС як з глюкозою, так і з сахарозою (рис. 3.3 б).

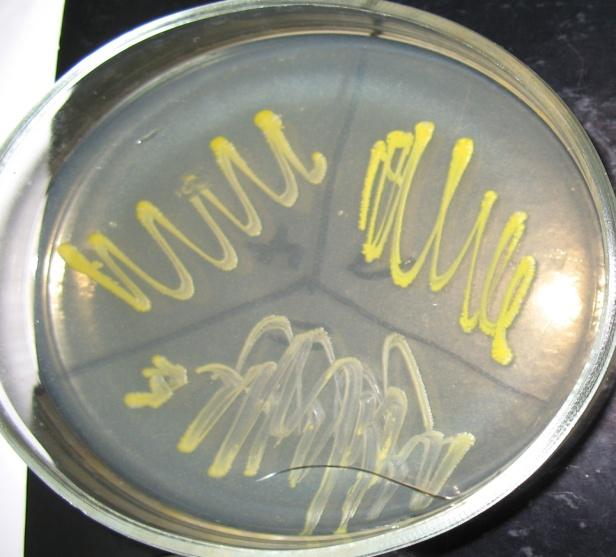
а


б
Рис. 3.3 Позитивний та негативний контроль дослідження ауксотрофності клонів Brevibacterium
Негативний контроль на середовищі Адамса не давав росту клонів № 2, 5, 7, 8 та 11. Винятком був клон №6, який не потребував факторів росту взагалі.
Для кожного клону проводили паралельні посіви на МС з амінокислотами. Джерелом вуглецевого живлення були глюкоза та сахароза. На середовищі з глюкозою більшість клонів не проявляли ауксотрофність і пігмент не утворювали. На рисунку 3.4 показано залежність росту клонів від лейцину та гомосерину (середовище з сахарозою).

а

б
Рис. 3.4 Ауксотрофність клонів до лейцину та гомосерину на МС з сахарозою
Бревібактерії утворювали пігмент на МС, проте проявляли ауксотрофність у відношенні більш широкого кола амінокислот (табл. 3.3).
Таблиця 3.3
Визначення ауксотрофності клонів Brevibacterium на твердому мс з сахарозою
|
Середовища, амінокислоти |
КЛОНИ | |||||
|
2 |
5 |
6 |
7 |
8 |
11 | |
|
Контроль ПС |
+++ |
+++ |
+++ |
+++ п |
+++ |
+++п |
|
Контроль МС |
- |
- |
- |
- |
- |
- |
|
Аспарагін |
- |
- |
- |
- |
- |
- |
|
Треонін |
- |
- |
++ |
- |
+ |
+ |
|
Ізолейцин |
- |
- |
+ |
- |
+ |
- |
|
Лейцин |
+++ |
+++ |
+++ |
+++ п |
+++ п |
+++п |
|
Тирозин |
- |
- |
- |
- |
+ |
- |
|
Метіонін |
- |
+ |
++ |
+ п |
- |
+ |
|
Триптофан |
+ |
+ |
+++ |
- |
- |
+ |
|
Гомосерин |
+ |
- |
+++ |
- |
- |
+ |
Примітка. +++ - інтенсивний ріст; + - наявність росту;– - відсутність росту; п– жовтий пігмент.
Як видно з таблиці 3.3 всі клони були лейцинозалежними і набули ауксотрофності до метіоніну (№ 5, 7, 11), треоніну (№8, 11), ізолейцину (№8), триптофану (№2, 5, 11) та гомосерину (№2, 11). Ріст клону №6 був позитивним на МС.
В подальших дослідженнях було здійснено культивування штамів-продуцентів на рідкому середовищі МС з амінокислотами тільки аспартатної родини (табл. 3.4). Серед всіх клонів було вибрано три найбільш продуктивних (клони №6, №7, №11) за лізином (див. табл. 3.2).
На рідкому середовищі МС з різними амінокислотами було підтверджено ауксотрофність за лейцином та метионіном клону № 7, який інтенсивно ріс, продукуючи пігмент, змінюючи рН середовища і синтезуючи лізин та інші амінокислоти.
У дослідах, де був відсутній ріст (незначна зміна ОГ), продуктивність за лізином не визначали. У дослідах з аспарагіном та ізолейцином при незначній зміні рН та ОГ продуктивність за лізином не визначали.
Таблиця 3.4