

Клітинна організація
Розміри клітин варіюють від 0,1-0,25 мкм.Органоіди еукаріотичних клітин : - в ядрі - хромосоми,
- в цитоплазмі – рибосоми , мітохондрії ,ендоплазматичний ретикул, комплекс гольджи, лізосоми, клітинна мембрана. Клітини мають мембранні структури, які підтримують форму – мікротрубочки, мікрофібрили, та вклюбчення. Білки – ферменти, вонисинтезуються лише клітині.
Особливіст клітини – компартменталізація (компартментація) – просторова організація хімічних процесів, дихання , на мембранах хромосом, синтез білку – на рибосомах.
Багатоядерність – збільшує концентрацію молекул матрични хРНК в цитоплазмі
Піноцетоз - захват з середовища капель з крупними молекулами – білки.
Фагоцитоз – захват круних частин, вірусі та літин.
Процес отримання клітинами стійкх внутрішніх відмінностей повязані з виконанням функцій – називається дифференцировкою. Вона повязана з синтезом білків. Наприклад синтез колагену.Люба клітина організму багатоклітинного
володіє однаковм повним фондом генетичного матеріалу та потенціями для роявленн генетичної інформації – тотипотентність .
Процес отримання клітинами стійкх внутрішніх відмінностей повязані з виконанням функцій – називається дифференцировкою. Вона повязана з синтезом білків. Наприклад синтез колагену.Люба клітина організму багатоклітинного
ДНК - дуже довгі молекули, вони локалізовані в ядрі та повязані з білами - гістонами вони включають некодуючі області – інтрони. ДНк прокаріот – короткі кільцеві молекули, які місяться в цитоплазмі вони не мають інтронів. Клітки еукаріот складаються з спеціалізованих відділів –органел.Поцеси снтезу та созрівання РНКі іблків у еукаріот протікають в різнгих відділах клітин та механізми їх регулювання не залежать один від одного У клітинах укаріот повний набір генетичної інформації знаходиься в молекулі ДНК, з яких сформовано лінейні структури – хромосоми, які заключені в ядро, а еукаріотичні клітини містяться в цитоплазмі. Цитопзазмою називають область клтин між ядром та плазматичною мембраною. Цитозоем називають усе містиме клітини за виключенням органел.
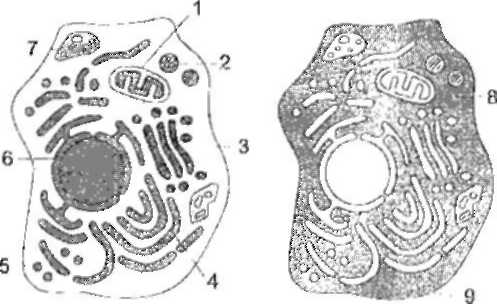
Рис- схема еукарітоитчної кліини : 1 – мітохондрії , 2 – пероксисома, 3 – апарат Гольджи, 4 – ендоплазматичний ретикул 5 – везикула , 6 – стінка ядра, 7- лізосома 8 – цитозоль 9 – плазматична мембрана.
Лекція 3.Компоненти клітинКЛІТИННА ОРГАНІЗАЦІЯ
Розміри клітин варіюють від 0,1-0,25 мкм.Органоіди еукаріотичних клітин : - в ядрі - хромосоми,
в цитоплазмі – рибосоми , мітохондрії ,ендоплазматичний ретикул, комплекс гольджи, лізосоми, клітинна мембрана. Клітини мають мембранні структури, які підтримують форму – мікротрубочки, мікрофібрили, та вклюбчення. Білки – ферменти, вонисинтезуються лише клітині.
Особливіст клітини – компартменталізація (компартментація) – просторова організація хімічних процесів, дихання , на мембранах хромосом, синтез білку – на рибосомах.
Багатоядерність – збільшує концентрацію молекул матрични хРНК в цитоплазмі
Піноцетоз = захват з середовища капель з крупними молекулами – білки.
Фагоцитоз – захват круних частин, вірусі та літин.
Процес отримання клітинами стійкх внутрішніх відмінностей повязані з виконанням функцій – називається дифференцировкою. Вона повязана з синтезом білків. Наприклад синтез колагену.Люба клітина організму багатоклітинного володіє однаковм повним фондом генетичного матеріалу та потенціями для роявленн генетичної інформації – тотипотентність .
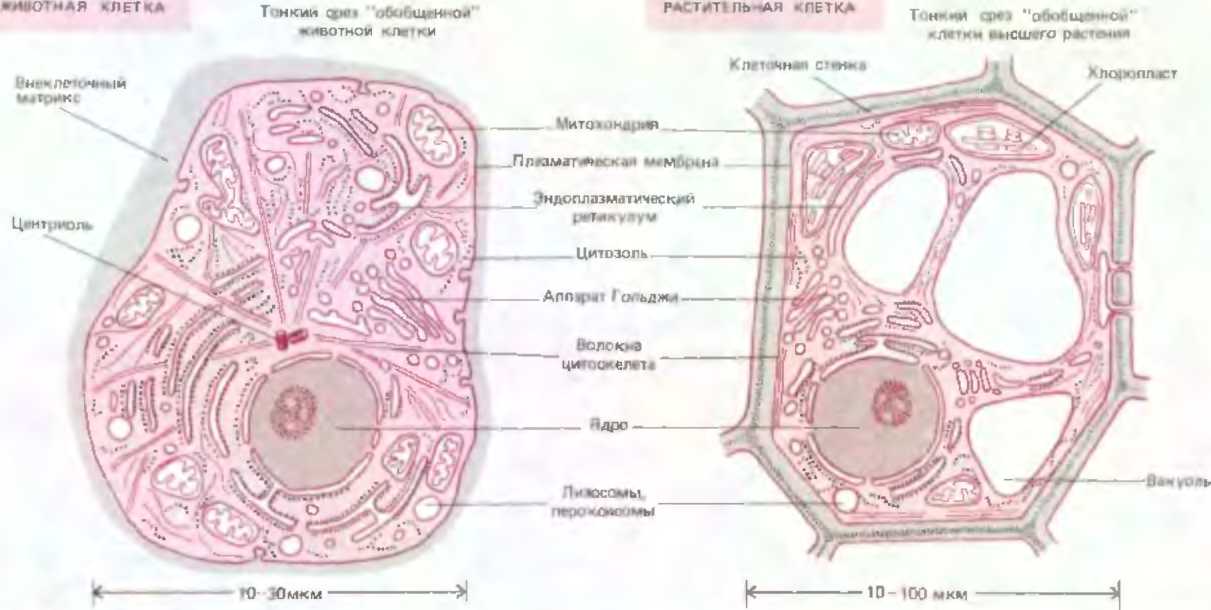
Рис. Будова рослиної та тваринної клітини
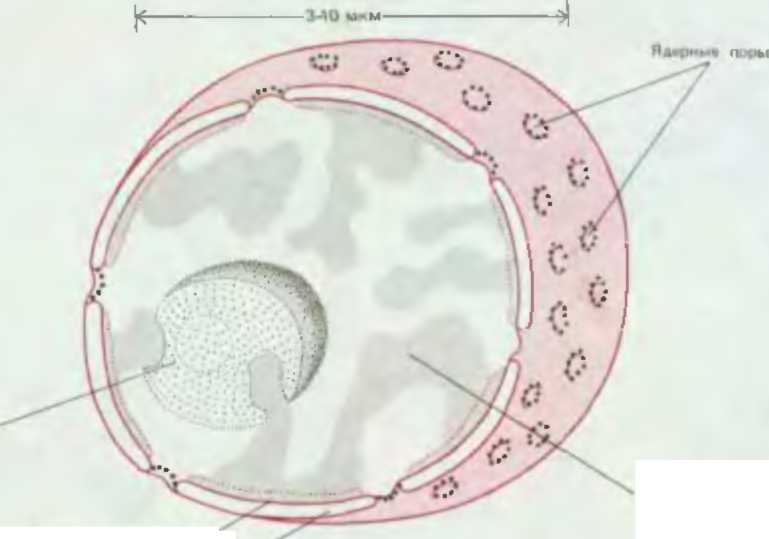
Хроматин
Клітини еукариот містять декілька характерних органел
Як же вплинуло накопичення молекулярного кисню в атмосфері на анаеробні організми, що поклали початок життя на Землі? У світі, багатому киснем, який не міг бути ними використаний, такі організми опинилися в невигідних умовах. Деякі з них, без сумніву, вимерли. Інші або розвинули здатність до дихання, або
Вона пронизана білковими вололкнами , які складають цитоскелет., Він містить оргпноїди та мембранну вакуолярну систему. Взаємодія між ними здійснюється через рідинну складову- цитозоль.- колоїдну систему, яка містить 75-85 % води 10-12 білків та амінокислот, 4-6,% вуглеводів, 2-3 % лпідів, 1 % інших. Цитозоль займає 50 % обєму клітини.В цитоплазмі є мітохондрії та рибосоми. Мітохондрії місять ферменти та та білково-мембранні структури для окислення орг.зєднань.та синтезу АТФ. Пластіди є у фотосинтезуючих та простійших.
Мітотохондрии і хлоропласти (мал. 1-19 і 1-20). Кожна з цих органел оточена власною подвійною мембраною, що відрізняється за хімічними властивостями від мембрани, що оточує ядро. Мітохондрії - майже універсальний компонент эукариотических клітин, тоді як хлоропласти зустрічаються лише в тих эукариотических клітинах, які здатні до фотосинтезу, т. е. у клітинах рослин, але не тварин і грибів. Вважається, що обоє органелльг мають симбіотичне походження.
1.2.6. Эукариотические клітини залежать від мітохондрій, осушествляюших окислювальний метаболізм Мітохондрії багато в чому схожі на свободноживушие прокариотические організми: наприклад, вони нагадують бактерій за формою і розміром, містять ДНК, виробляють білок і розмножуються діленням. Зруйнувавши эукариотические клітини і розділивши їх компоненти, можна показати, що мітохондрії відповідальні за дихання і що ні вяких частинах клітин цей процес не відбувається. Без мітохондрій клітини тварині грибів були б анаеробами, залежними у своїх енергетичних потребах від порівняно малоефективного і архаїчного процесу гліколізу. Багато сучасних бактерій можуть дихати, причому механізм цього дихання має явну схожість з диханням у мітохондрій, і є усі підстави думати, що эукариотические клітини є нащадками примітивних анаеробних організмів, які вижили у багатому киснем світі, поглинувши аеробних бактерій. Вони підтримували їх в стані симбіозу заради властивої їм здатності споживати атмосферний кисень і виробляти енергію. Аналіз деяких сучасних організмів свідчить про можливість такої еволюційної події. Існує декілька сотень видів одноклітинних эукариот, які нагадують гіпотетичний предковый эукариотический організм тим що живуть в умовах дефіциту кисню (наприклад, в кишковику тварин) і зовсім не мають мітохондрій. Порівняння нуклеотидных послідовностей, проведене останнім часом, дає основу припускати, що одна група таких організмів, мікроспоридії, дуже рано в еволюції відгалузилася від лінії, що веде до інших эукариотическим клітин Інший представник эукариот. амеба Pelomvxa palustris, хоча і позбавлений мітохондрій, все ж здійснює окислювальний метаболізм, "прихистивши" у своїй цитоплазмі аеробних бактерії і встановивши з ними постійні симбіотичні стосунки. Таким чином, існування мікроспоридій і Pelomyxa свідчить про реальність двох етапів в еволюції эукариот, до яких відносимося і ми самі.Придбання мітохондрій повинне було мати багато наслідків. Наприклад, у прокариотических клітин плазматична мембрана тісно пов'язана з утворенням енергії, тоді як у эукариотических клітин ця найважливіша функція передана мітохондріям. Здається цілком вірогідним, що звільнення плазматичної мембрани эукариотической клітини від цієї функції дозволило їй набути нових властивостей Зокрема, оскільки эукариотическим клітинам не треба підтримувати високий градієнт Н+ на своїй мембрані (що потрібне для виробництва АТР у прокариот), у них з'являється можливість використовувати контрольовані зміни в іонній проникності плазматичної мембрани в цілях клітинної сигналізації. Отже, одночасно з виникненням эукариот в плазматичній мембрані з'являються іонні канали. Нині у вищих організмів ці канали опосередкують складні процеси передачі електричних сигналів (зокрема, в нервовій системі), а у одноклітинних вільноживучих эукариот, таких, як прості, вони багато в чому визначають їх поведінку (див. нижче).
Хлоропласти здійснюють фотосинтез значною мірою так само, як прокариоты-цианобактерии, сонячне світло у них поглошается приєднаним до мембран хлорофілом. Деякі Хлоропласти по будові багато в чому нагадують ціанобактерії наприклад схожими можуть бути їх розміри і спосіб укладання в шари хлорофиллсодержащих мембран (мал. 1-20). Показано також, що хлоропласти розмножуються діленням, а нуклеотидная послідовність їх ДНК майже повністю гомологична певним ділянкам бактерійної хромосоми. Усе це наводить на думку, що хлоропласти і ціанобактерії мають загального предка і що хлоропласти пішли від прокариот, "захоплених" колись эукариотическими клітинами. Прокариоты здійснювали фотосинтез для клітин-хазяїв в обмін на ті, що надаютьсяНа мал. 1-22 показано еволюційне походження эукариот відповідно до симбіотичної теорії Слід зазначити, що мітохондрії і хлоропласти, проявляючи певну схожість з сучасними аеробними бактеріями і ціанобактеріями, в той же час у багатьох відношеннях відрізняються від них. Наприклад, кількість ДНК в цих органелах дуже мало, більшість складових їх молекул синтезуються поза органелами і лише потім в них гранспортируется. Якщо вважати, що мітохондрії і хлоропласти дійсно виникли з симбіотичних бактерій, то слід визнати, що вони зазнали значні еволюційні зміни і стали дуже залежними від своїх хазяїв.
Для сучасних эукариот характерно не лише наявність мітохондрій, їм властивий цілий ряд особливостей, що відрізняють їх від прокариот (таблиця. 1-1). Разом усі ці особливості наділяють эукариотические клітини великою кількістю різних потенційних можливостей, і важко сказати, яка з них виникла раніше інших. Помітимо, що найважливішим кроком на шляху еволюції була поява мітохондрій в анаеробних эукариотических клітинах, оскільки разом з ними клітини отримували ефективне джерело енергії і могли направити її на ускладнення своїх функцій.
Об'єм эукариотических клітин, як правило, в 1000 і більше разів перевищує об'єм клітин прокариот. Відповідно більше в эукариотических клітинах і різноманітного клітинного матеріалу, наприклад зміст ДНК в клітинах людини в 1000 разів перевищує її кількість в клітинах бактерій. Відомо, що саме на мембрані протікає ряд найважливіших реакцій, пов'язаних зі вступом в клітину сировини для метаболізму і виходом відповідних продуктів в позаклітинний простір. Ось чому великий об'єм эукариотических клітин вимагає значного збільшення поверхні їх мембрани в порівнянні з клітинами прокариот. Але згідно із законами геометрії, при простому збільшенні розмірів якого-небудь предмета його об'єм зростає як куб лінійного розміру, а плошадь поверхні - лише як квадрат Тому для збереження необхідного співвідношення площі поверхні і об'єму великі эукариотические клітини вимушені збільшувати свою поверхню за рахунок вигинів, складок і інших ускладнень форми мембрани.
1.2.8. Эукариотические клітини містять безліч різних внутрішніх мембран.Складність будови мембран - одна з основних особливостей усіх эукариотических клітин. Мембрани оточують ядро, мітохондрії іендоплазматичний ретикулум (ЕР). Ділянки гладкого ЕР беруть участь в ліпідному обміні, ділянки шорсткого, усипані рибосомами, є місцями синтезу білків. Синтезовані білки покидають цитозоль і входять в деякі інші компартменты клітини.
-
Обозначения
Амінокислоти
Ашто ас1
рус
сим п.
ащ'л.
Заменимые аминокислоты
I
Алании
2
Аспаргин
3
Аспаргиновая кислота
4
Глицин
5
Глутамип
6
Глутаминовая кислота
7
Пролин
8
Серии
9
Тирозин
10
Цистеии
Частично заменимая аминокислота
11
Аршнин
Незаменимые апиокислоты
12
Валин
13
Гист идин
14
Изолейцин
15
Лейцин
16
Лизин
17
Метионин
18
Треонин
19
Триптофан
20
Фенилаланин
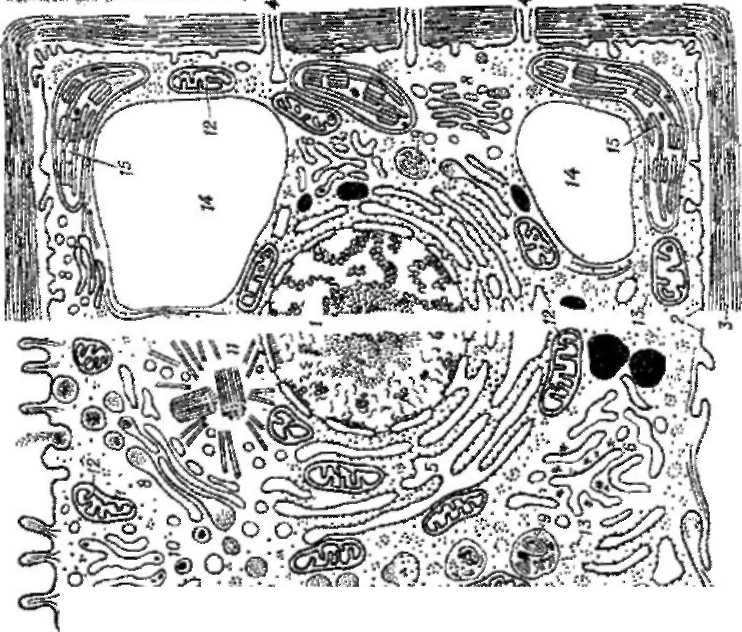
Мал. 1-24. Електронна мікрофотографія зрізу клітини ссавця. Видно апарат Гольджи. Ця структура складається з декількох шарів сплощених мембранних бульбашок (см також схему 1-1). Апарат Гольджи бере участь в синтезі і упаковці матеріалу, призначеного для секреції з клітини, а також в транспорті новосинтезованих білків у відведений для них клітинний компартмент. (З люб'язного дозвол рослин) хлоропласти. Вони утворюють лабіринт ендоплазматичного ретикулума (рис L - 23), де синтезуються ліпіди і мембранні білки, а також матеріал, призначений для експорту з клітини. Мембрани формують стопки сплощених бульбашок, що становлять апарат Гольджи (мал. 1-24). який теж бере участь в синтезі і транспорті різних органічних молекул Мембрани оточують лізосоми, що містять запас ферментів, необхідних для внутрішньоклітинного травлення, і таким чином захищають від дії цих ферментів білки і нуклеїнові кислоти самої клітини. Так само мембрани оточують пероксисоми, де в процесі окислення різних молекул утворюються і розкладаються небезпечні высоко-реакционноспособные перекиси (пероксиды). Мембрани утворюють також маленькі везикули (бульбашки) і великі, заповнені рідиною вакуоли (у рослин). Усі ці оточені мембранами структури відповідають визначеним компартментам цитоплазми. В сукупності вони займають майже половину об'єму типової живої клітини. Цитоплазматичний компартмент, позбавлений усіх оточених мембранами органел, зазвичай називають цитозолем.Усі перераховані нами мембранні структури знаходяться усередині клітини. Як же у такому разі вони можуть впливати на площу її поверхні? Відповідь полягає в тому, що межд> внутрішньоклітинними, оточеними мембранами структурами і позаклітинним середовищем відбувається обмін. Він здійснюється за допомогою двох унікальних для эукариотических клітин процесів: эндоцитоза і екзоцитозу. При эндоцитозе деякі ділянки зовнішньої поверхневої мембрани впячиваются і відриваються, утворюючи цитоплазматичні мембранні бульбашки, що містять речовини, які знаходилися в зовнішньому середовищі або були адсорбовані на поверхні клітини. У такий спосіб в цитоплазму можуть потрапити дуже великі частки і навіть цілі клітини (фагоцитоз) Екзоцитоз – зворотний процес, при якому внутрішньоклітинні мембранні бульбашки зливаються з плазматичною мембраною, вивільняючи тим самим свій вміст в зовнішнє середовище. Завдяки описаному механізму, розташовані глибоко усередині клітини і оточені мембранами компартменты збільшують ефективну поверхню клітини, що бере участь в обміні речовин із зовнішнім середовищем.Як ми побачимо в подальших главах, різні мембрани і оточені ними компартменти эукариотических клітин стали високоспеціалізованими: одні призначені для секреції, інші – для всмоктування, треті - для певних біосинтетичних процесів ит. д.