

Мацерация. Межклетники
Общая для двух соседних клеток склеивающая их срединная пластинка в процессе онтогенеза может разрушаться, клетки при этом разъединяются. Этот процесс носит название мацерации (лат. maceratio, от macero - размягчаю, размачиваю), он связан с растворением пектиновых веществ срединной пластинки ферментом пектиназа с последующим вымыванием продуктов распада. У растений в естественных условиях мацерация происходит в мякоти созревающих сочных плодов (яблок, груш, томатов, бананов, персика, дыни и т. д.). Запасающая ткань плодов при этом распадается на отдельные клетки, зрелые (и особенно перезрелые) плоды имеют мучнистый характер. Этот же процесс происходит при раскрывании сухих плодов (коробочка мака, бобы гороха, фасоли, стручки крестоцветных и т. д.), в отделительном слое перед опадением листьев, в плодоножках и лепестках перед их опадением.
Искусственная мацерация (с помощью микроорганизмов) широко используется в промышленности, например при обработке (мочке) волокнистых растений (лен, конопля).
У растений с возрастом наблюдается частичная мацерация клеток, при которой срединная пластинка растворяется не полностью, а участками, чаще всего по углам. Под действием тургорного давления клетки округляются и между ними образуются полости - межклетники. Постепенно они увеличиваются в размерах, из трех-, четырехугольных узких щелей превращаются в цилиндрические или многогранные каналы, вытянутые вдоль продольной оси органа.
Частичное разрушение срединной пластинки - не единственный путь образования межклетников. В общем они могут формироваться в процессе гистогенеза как при раздвижении соседних клеток - схизогенные межклетники, так и при их разрыве и последующем отмирании - рек-сигенные или при растворении группы клеток - лизигенные. Возможно также их смешанное образование. Сливаясь друг с другом и образуя единую систему каналов, сообщающуюся с внешней средой через устьица, межклетники улучшают газообмен в клетках, могут быть вместилищами разнообразных продуктов выделения (слизей, камедей, смол, млечного сока, эфирных масел и т. д.); в них поступают пары воды при транспирации. Многие растения (водные, заболоченных участков и т. д.) содержат паренхимную ткань с крупными межклетниками, наполненными воздухом (см. Аэренхима).
Межклеточные связи
Процесс мацерации убедительно доказывает, что каждая клетка окружена собственной оболочкой и является самостоятельной структурной единицей в многоклеточном теле высшего растения (и самостоятельным одноклеточным организмом на низших ступенях эволюции). Однако обособленность клеток не означает их полную изолированность друг от друга. Клетки сообщаются между собой за счет структур дифференцирующейся клеточной оболочки - плазмодесм, пор и перфораций.
Плазмодесмы (греч. плазма и desmos - лента, связь). Представляют собой цитоплазматические нити, соединяющие протопласты соседних растительных клеток (рис. 66). Впервые описаны в 1877 г. русским ученым И. Н. Горожанкиным, затем Э. Танглем (1879), а свое название получили от Э. Страсбургера (1882).
Рис. 66. Плазмодесмы на продольном срезе меристемы корня кукурузы (Zea mays): 1 - замыкающая пленка поры; 2 - плазмодесмы; 3 - срединная пластинка |
Диаметр плазмодесм лежит на границе разрешения светового микроскопа, поэтому наблюдать их без особой предварительной обработки, вызывающей разбухание оболочки, можно очень редко, например в запасающей ткани семян пальмы, хурмы, конского каштана. Электронно-микроскопические исследования позволили обнаружить плазмодесмы почти у всех растений и во всех тканях. Количество их варьирует в разных клетках и растениях в пределах нескольких сот и даже тысяч (например, в клетках кончика лука - до 10 000-20 000 на одну клетку, в паренхимных клетках омелы – 6000-24 000 и т. д.). Поперечник плазмодесмы составляет 20-70 нм (чаще 30-40 нм). Образование плазмодесм начинается в период заложения клеточной пластинки: между сливающимися пектиновыми вакуолями фрагмопласта остаются перешейки, где цитоплазма между дочерними клетками не разъединяется. Эти перешейки и представляют собой будущие плазмодесмы (см. рис. 66). В период образования клеточной пластинки, а затем и первичной оболочки будущие плазмодесмы, расположенные группами, приурочены в основном к замыкающим пленкам пор (см. дальше). Располагаются в канальцах, полость которых выстлана наружной мембраной плазмодесм - плазмалеммой, чем достигается непрерывность цитоплазмы всего организма. |

Через них проходит по меньшей мере один трубчатый тяж эндоплазматического ретикулума - десмотрубочка, непосредственно переходящая в цистерны ретикулума, расположенные напротив отверстий плазмодесм. Остальная часть канала заполнена матриксом цитоплазмы. При образовании вторичной оболочки плазмодесмы, располагающиеся группами, объединяются в виде поровых полей, но только в живых клетках, оболочки мертвых клеток плазмодесм не несут.
Таким образом, плазмодесмы связывают между собой протопласты различных клеток в единое целое - симпласт. С помощью плазмодесм различные вещества передаются в виде растворов от клетки к клетке, происходит передача раздражений и гормонов. Вероятно, таким путем в различные ткани проникают вирусы. Транспорт веществ через плазмодесмы получил название сим-пластического в отличие от апопластического транспорта по оболочкам и межклетникам.
Поры (греч. poros - отверстие). В фазе растяжения на всех стенках первичной оболочки можно видеть участки, пронизанные плазмодесмами, в которых отложение оболочки происходит менее интенсивно. В связи с этим появляются многочисленные углубления различных размеров, которые и называются первичными норовыми полями (рис. 67, А). Их бывает очень много, особенно в оболочках эмбриональных клеток, иногда целиком одна из стенок превращается в поровое поле.
При вторичном утолщении оболочки отложение ее слоев, как правило, не происходит в участке первичных поровых полей, а нередко и в других точках. Возникают неутолщенные места (прорывы) вторичной оболочки, которые называются порами (рис. 67, Б). Следовательно, поры характерны только для клеток со вторичной оболочкой. Если она слабо развита, поры представлены мелкими углублениями, в типичном случае - имеют вид трубчато радиальных каналов, тянущихся через мощную вторичную оболочку от полости клетки к первичной стенке. Их можно хорошо рассмотреть у каменистых клеток (см. Механические ткани), находящихся во многих плодах (рис. 67, В).
Поры двух смежных клеток обычно располагаются друг против друга - пары пор. Каждая пора, входящая в состав данной пары, имеет поровую камеру, обе камеры отделяются друг от друга тонким участком оболочки — замыкающей пленкой поры, или поровой мембраной, состоящей из двух первичных оболочек и срединной пластинки (рис. 67, Б).
При формировании пор в замыкающей пленке сохраняются плазмодесмы, обеспечивающие связь между цитоплазмой, заполняющей камеры пор двух соседних клеток. По мере утолщения вторичной оболочки камеры превращаются в каналы. Уменьшение внутренней окружности растущей оболочки может вызвать слияние соседних поровых каналов, образуются так называемые ветвистые поры (рис. 67, В). Если клетка теряет живое содержимое в процессе дифференциации, плазмодесмы и цитоплазма в камерах исчезают.
Поры чаще встречаются в стенках оболочек, разделяющих соседние клетки. Реже они встречаются в наружных, граничащих с атмосферой оболочках клеток стебля и листьев (некоторые злаки, лианы).
|
Рис. 67. Поры: А - первичные поровые поля (схемах); Б - образование пары пор (схемат.); В - простые поры в каменистых клетках плодов айвы (Cydonia oblonga); Г - две окаймленные пары пор (а - вид сбоку, б - вид окаймленной поры с поверхности; в - закрытая окаймленная пора); Д - окаймленная пора в трахеитах весенней древесины сосны (Pious pungens); 1 - первичная оболочка; 2 - срединная пластинка; 3 - первичное поровое поле с плазмодесмами; 4 - апертура пор; 5 - замыкающая пленка поры; 6 - вторичная оболочка; 7 - ветвистые поры; 8 - полость клетки; 9 - торус; 10 - маргинальная зона; 11 - окаймление; 12 - полость поры |
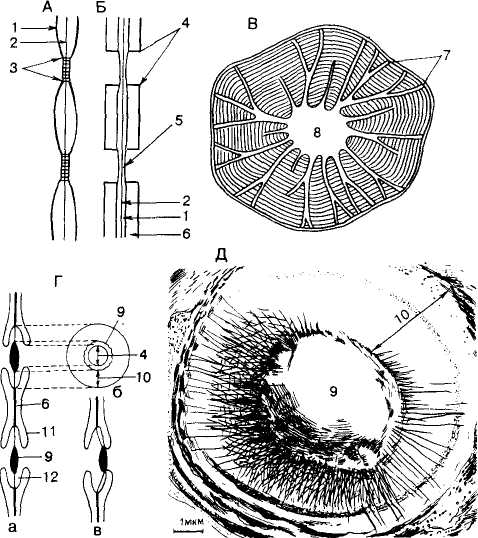
Поры различают по размерам и тонкой структуре. Вторичная оболочка может резко прерываться у краев камеры поры, диаметр которой благодаря этому почти не изменяется по всей толще вторичной оболочки. Поры этого типа называются простыми, а комбинация двух простых пор - простой парой пор (см. рис. 67, В). Вторичная оболочка может нависать над камерой поры в виде свода, образуя ее окаймление. Такие поры получили название окаймленных или окаймленной пары пор (рис. 67, Г, Д). Поровая камера, ограниченная окаймлением, открывается в полость клетки через отверстие в окаймлении - апертуру поры.
В трахеидах голосеменных растений, главным образом у представителей сем. сосновых (Pinaceae), замыкающая пленка, разделяющая окаймленную пару пор, имеет высокоспециализированную структуру (рис. 67, Г). Утолщение в средней части замыкающей пленки образует торус, остальная часть - краевая, или маргинальная, зона - состоит из радиально отходящих от торуса пучков микрофибрилл. Замыкающая пленка в маргинальной зоне эластична и при давлении на нее содержимого трахеид смещается к одной или другой стороне окаймления, закрывая с помощью торуса апертуру поры. В таком состоянии пора называется закрытой, не функционирующей в проведении веществ (рис. 67, Г, В).
Если вторичная оболочка очень мощная, окаймленная пора утолщается. Довольно маленькая камера такой поры соединяется с полостью клетки узким каналом поры, который у многих пор напоминает сжатую воронку. Наружная апертура (отверстие), обращенная к полости клетки, небольшая и круглая, а внутренняя - длинная и щелевидная (рис. 67, Г).
У листопадных пород деревьев замыкающая пленка окаймленных пор лишена торуса и обладает такой же текстурой, как и первичная оболочка.
По типу расположения окаймленных пор различают клетки с лестничной (щелевидные поры располагаются в один ряд наподобие ступенек), супротивной (мелкие многочисленные поры расположены в горизонтальных рядах) и очередной (поры расположены в шахматном порядке) поро-востью (см. Сосуды).
Поры играют важную роль в проведении воды и питательных веществ по всем тканям растений, являясь своего рода микрофильтром благодаря тонкости замыкающей пленки. Поры имеются в клеточной оболочке и низших растений.
Перфорации (лат. perforatio - пробуравливание, проникновение). Представляют собой сквозные отверстия (дырки) в клеточных оболочках. Встречаются преимущественно в клеточных оболочках сосудов и ситовидных трубок растений. В сосудах перфорации возникают на месте окаймленных пор в оболочках между смежными проводящими элементами. Они могут быть множественными (например, лестничными — в сосудах папоротников), эфедроидными (состоят из группы округлых отверстий у гнетовьгх) и простыми (одно общее отверстие на поперечных стенках члеников сосудов у многих цветковых растений), По перфорациям свободно продвигаются питательные растворы. Подробнее образование в структура перфораций рассматриваются в разделе «Проводящие ткани».
Заканчивая изложение главы «Растительная клетка», следует подчеркнуть, что именно на клеточном и тканевом уровнях организации, изучаемых структурной ботаникой, складываются специфические, далеко еще не понятые свойства живых организмов, познание которых и является важнейшей предпосылкой для управления развитием растении.
В литературе последних лет (1987-1996)* отмечается, что бурного расцвета структурная ботаника сейчас не переживает. * Васильев А Е. Состояние и перспективы развития структурной ботаники (цитологии, анатомии, эмбриологии, морфологии) // Бот. журн. 1989. Т. 74. № 8. С. 1090-1100. И все же определенные достижения и даже открытия в этой области за последние годы были и предвидятся в будущем, и наиболее серьезные - в цитологии (морфологии клетки).
Из открытий в области цитологии следует назвать обоснование представления о полуавтономности не только пластид и митохондрий, но и о симбиотическом происхождении пероксисом, подобно митохондриям и пластидам.
Установлено, что плазмодесмы, которые, как всегда считалось, представляют мостики цитоплазмы между двумя дочерними клетками, оставшиеся в разделившей их перегородке при цитокинезе, могут возникать и между недочерним клетками и даже между клетками разных видов и родов растений - при прививках, у химер (грет Chimaira - мифическое чудовище с головой льва, туловищем козы и хвостом дракона; организм-мозаик, который сочетает в себе клетки, ткани и органы разных организмов), в зоне контакта клеток паразита и растения-хозяина. Встает вопрос, как может цитоплазма каждой смежной клетки «пробуравить» в толстой прочной оболочке навстречу друг другу тончайшие отверстия с идеальным отверстием в центре, каков механизм образования этих многочисленных структур? Получены данные, что диаметр плазмодесм на входа может обратимо меняться, при этом регулируете) скорость тока веществ между клетками.
При исследовании клеточной оболочки важные достижения получены при помощи комплекса современных методов. Сформулировано представление о геликоидной текстуре микрофибрилл целлюлозы в оболочке, согласно которому фибриллы откладываются по спирали в виде многочисленных очень узких слоев, причем в соседит слоях происходит изменение спирали на небольшой угол, в результате во всей оболочке слои как бы вращаются. Неясно, как такие многослойные оболочки растут растяжением.
На плазмалемме открыты особые мультиферментные целлюлозосинтезирующие комплексы в виде розеток. Предполагают, что они плывут в плоскости мембраны, оставляя после себя готовые микрофибриллы. Так осуществляются синтез целлюлозы, кристаллизация и ориентация фибрилл.
Чрезвычайно актуальным остается изучение клеточной оболочки и прежде всего механизма ее образования. Пока не удается воспроизвести искусственно формирование микрофибрилл целлюлозы. Познание этого процесса позволит искусственно получать целлюлозу и отказаться от синтетических волокон.
Важнейшим направлением в области морфологии клетки в настоящее время является исследование цитоскелета - опорно-двигательного аппарата клетки, изучаемого с помощью комплекса методов. Уже выявлена огромная роль компонентов цитоскелета, микротрубочек, микро-филаментов, микротрабекул, в ряде жизненно важных процессов - движении цитоплазмы и независимом движении отдельных органоидов, определении формы клетки, ориентации целлю-лозосинтезирующих комплексов и образуемых ими микрофибрилл целлюлозы, в движении хромосом в период деления ядра.
Другое важное направление в цитологии растений - объемная реконструкция структурно-функциональных компонентов клетки, в особенности аппарата Гольджи и эндоплазматического ретикулума по серии ультратонких и полутонких срезов с помощью компьютера. Это поможет познать взаимодействие органоидов в ряде процессов жизнедеятельности клетки.
Определенный интерес представляют исследования вакуолей и органоидов самопереваривания цитоплазмы - лизосом. Неясны пока вопросы происхождения вакуолей, их рост, связи с системой лизосом. Весьма вероятно, что, подобно пластидам, митохондриям, пероксисомам, вакуоли самовоспроизводятся.
В настоящее время в цитологии преобладают молекулярная биология клетки, клеточная физиология и биохимическая цитология. Данные цитологии широко используются в разработке проблем происхождения эукариотной клетки, происхождения растений.