

6.3. Сложные липиды
К сложным липидам относятся гликолипиды, фосфолипиды (фосфатиды), сфинголипиды.
Гликолипиды. Гликолипиды — сложные соединения, содержащие липидный и углеводный фрагменты, соединенные ковалент- ной связью. В последние годы интерес к этим соединениям возрос, так как они выполняют не только структурные, но и регуляторные функции. Они входят в состав плазматических мембран и мембранных систем клетки, обладают антигенными свойствами, определяют серологический тип бактерий. Гликолипиды были выделены из растений (зерно пшеницы, водоросли, зеленые листья растений), из животных тканей (головной мозг), из микроорганизмов; углеводным компонентом их является галактоза (например, моногалактозилдиацилглицерин):
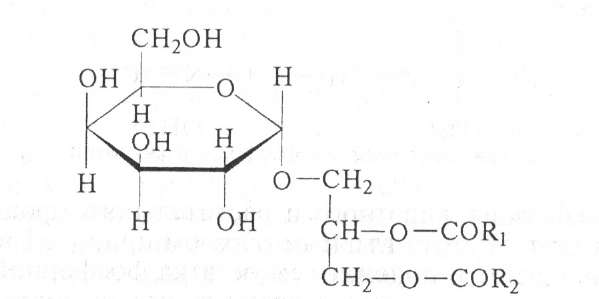
где R1 и R2 — остатки жирных кислот. В составе нейтральных гли- колипидов в значительных количествах присутствуют остатки ненасыщенных жирных кислот (олеиновая, линолевая и др.). Основным углеводным компонентом кислотоустойчивых бактерий являются: D-арабиноза, D-глюкоза, D-манноза, D-рамноза, О-метилированные дезоксигексозы, трегалоза; липидный компонент представлен высокомолекулярными разветвленными жирными кислотами.
Фосфолипиды (фосфатиды). Фосфолипиды содержатся во всех тканях животных и растений. Много их в нервной ткани и ткани головного мозга животных, а также в желтке яиц птиц. Они содержат остатки глицерина, жирных кислот, фосфорной кислоты, азотистых оснований.
Различают: холинфосфатиды, коламинфосфатиды, серинфосфатиды, ацетальфосфатиды, инозитфосфатиды, сфингофосфатиды.
Фосфатиды нерастворимы в воде, но способны набухать в ней и образовывать водные эмульсии; хорошо растворяются в эфире, бензоле, хлороформе; в отличие от нейтральных жиров и стеринов нерастворимы в ацетоне (этим свойством пользуются для отделения фосфатидов от стеринов и жиров); легко окисляются благодаря наличию ненасыщенных жирных кислот.
Рассмотрим наиболее значимые фосфолипиды.
В качестве азотистого компонента фосфолипиды чаще всего содержат этаноламин, холин, инозит.
Холинфосфатиды, или лецитины, широко распространены в тканях животных, растений, микробов, входят в состав большинства мембран животных клеток. Много лецитина в ткани мозга, надпочечниках, эритроцитах. В яичном желтке его до 8...10%.
Холинфосфатиды образованы путем взаимодействия глицерина с двумя молекулами жирных кислот и молекулой фосфорной кислоты, которая соединена эфирной связью с азотистым основанием холином:

Холинфосфатиды животного и растительного происхождения отличаются друг от друга главным образом природой входящих в них жирных кислот и положением остатка фосфорной кислоты. В зависимости от положения последнего различаются а-лецитины, если остаток Н3Р04 расположен у 1-го углеродного атома глицерина, и р-лецитины, если в образовании эфирной связи с фосфорной кислотой участвует вторичный спиртовый гидроксил глицерина.
Как видно из формулы, в молекуле лецитина имеется кислотный гидроксил в остатке фосфорной кислоты и основный в остатке холина, поэтому лецитин может существовать в виде внутренней соли. Правильнее изображать формулу лецитина следующим образом:
В
состав лецитинов могут входить
олеиновая, пальмитиновая, линолевая,
арахидоновая и другие кислоты. При
гидролизе леци
тинов получают жирные кислоты, холин,
глицерофосфорную кислоту, которая
затем может распадаться на свободную
фосфорную кислоту и глицерин.

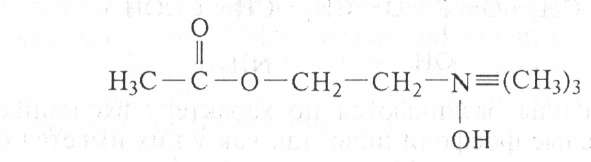
Ацетилхолин имеет важное значение в процессах нервной деятельности как переносчик нервного возбуждения (медиатор). Холин легко окисляется, превращаясь сначала в альдегид мускарин (впервые найден в грибе мухоморе), затем в кислоту бетаин, встречающуюся в тканях животных и растений.
Коламинфосфатиды (кефалины) построены так же, как и холинфосфатиды, из глицерина, жирных кислот и фосфорной кислоты, но в качестве азотистого основания в них входит этаноламин (коламин): CH2(NH2)CH2OH.
Кефалины широко распространены в природе. Впервые были найдены в ткани головного мозга (от лат. cephalus — голова), находятся в печени, почках, яичном желтке и др. Они растворимы в спирте и этим отличаются от лецитинов:

Серинфосфатиды по структуре аналогичны лецитину и кефалину, но в качестве азотистого соединения в них входит аминокислота серин:

Серинфосфатиды впервые были найдены в ткани мозга, имеют следующее строение:

Серинфосфатиды различаются по характеру входящих в них кислот. Это кислые фосфолипиды, так как у них имеется свободная карбоксильная группа. Между холинфосфатидами, коламинфосфатидами и серинфосфатидами существует родство: азотистые соединения этих фосфатидов могут переходить друг в друга:

Превращение серина в коламин происходит путем декарбокси- лирования, а коламина в холин — путем метилирования.
Инозитфосфатиды
имеют следующее строение:
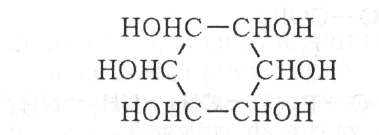
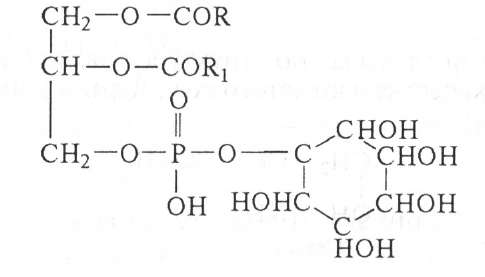
Впервые выделены из туберкулезных бацилл, а позже из растительных и животных тканей. Кроме обычных компонентов (глицерин, инозит, фосфорная кислота, жирные кислоты) в состав сложных инозитфосфатидов могут входить амины, аминокислоты, углеводные остатки.
Гликофосфоглицериды — углеводсодержагцие фосфолипиды — обнаружены в различных тканях. В них в качестве полярной группы выступает молекула углевода (их называют также фосфатидилсахарами). Связь между фосфолипидом и углеводом может быть О-эфирной или N-гликозидной. Так, выделены производные фосфатидилглицерина, содержащие остаток глюкозамина в 3-м положении (О-эфирная связь):

Фосфоглицериды, содержащие группу —ОСН=СН— (альдегидогенные), или фосфорсодержащие плазмогены, присутствуют во всех тканях животного организма. Их особенно много в головном и спинном мозге, сердечной мышце, где они входят в состав клеточных мембран.
Фосфолипиды составляют 25...30 % сухой массы мозга, из которых на долю плазмогенов приходится до 90 %.
Сфинголипиды. Это сложные эфиры сфингозина, входят в состав нервных клеток, тканей мозга, мембран эритроцитов, плазмы крови, печени и других органов и тканей; служат вторичными посредниками в передаче различных внешних сигналов в клетку; регулируют рост, дифференцировку, активацию и пролиферацию лимфоцитов, апоптоз клеток.
Сфингозин — алифатический двухатомный аминоспирт из 18 углеродных атомов:
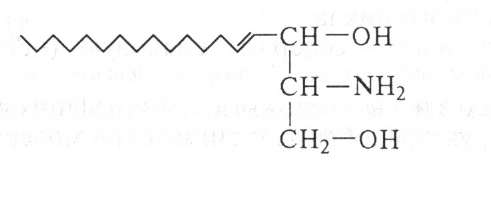
Сфингозин мобилизует ионы кальция из внутриклеточного депо и контролирует специфический Са+2-канал, влияет на синтез ДНК, конкурируя с гистонами.
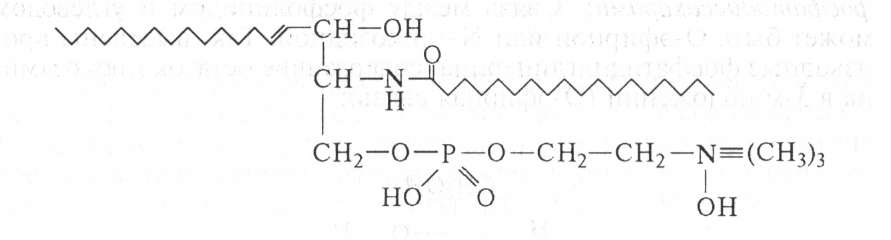
Сфингомиелин образуется из сфингозина путем замещения атома водорода аминной группы жирной кислотой и соединения одной гидроксильной группы через фосфорную кислоту с холином. Входит в состав нервных клеток:
Цероброзиды состоят из сфингозиновых оснований, жирной кислоты и гексозы (галактозы). Жирные кислоты их содержат 24 атома углерода, чаще всего встречаются невроновая, оксиневроновая, цереброновая кислоты:

Сульфатиды отличаются от цереброзидов наличием остатка серной кислоты. Они обладают кислым характером, играют важную роль в ионном транспорте через мембраны.
Церамидогексозы в качестве углеводного компонента содержат олигосахара (Р-лактоза). Регулируют рост и развитие нервных клеток, рост аксонов нейронов гиппокампа, играют роль в апоптозе, дифференцировке и старении клеток, а также в торможении клеточного цикла.
Гематозиды — содержат сиаловую (нейраминовую) кислоту.
Ганглиозиды — сложные гликосфинголипиды, разнообразны по структуре, образуют гигантские молекулы (до 400 моно меров), принимают участие в передаче нервного импульса:
