

Біологічна та біоорганічна хімія_Мардашко О.О._ изд. 2008-342 с._ОНМедУ-2012
.pdf
|
білком — білком-трансдуктором (від англ. trans- |
4. Секреція інсуліну острівцевими клітинами |
|||||||||||
|
duction — перенесення). |
|
|
|
підшлункової залози. |
||||||||
|
Тип D (кальцій/фосфатидилінозитолова |
5. Секреція адреналіну. |
|||||||||||
|
система) |
|
|
|
|
|
|
|
|
|
6. Секреція гістаміну мастоцитами. |
||
|
Багато рецепторів, зв’язуючись із гормоном чи |
|
|||||||||||
|
медіатором, активують мембранозв’язану фосфо- |
Механізм регуляції метаболізму |
|||||||||||
|
діестеразу — фосфоліпазу С. Активована фосфо- |
за допомогою іонів Са2+ |
|||||||||||
|
ліпаза С розщеплює фосфатидилінозитол 4,5-ди- |
Внутрішньоклітинний вміст іонів Са2+ дорів- |
|||||||||||
|
фосфат (похідне фосфоліпідів мембран), при цьо- |
||||||||||||
|
нює 10-7 ммоль/л, тоді як поза клітиною — |
||||||||||||
|
му виділяється інозитол-1,4,5-трифосфат і 1,2-ді- |
||||||||||||
|
ацилгліцерол. Ці сполуки є вторинними месенд- |
10-3 ммоль/л. Іони Са2+ надходять із зовнішнього |
|||||||||||
|
жерами і виявляють синергічну дію. |
середовища по «кальцієвих каналах» у мемб- |
|||||||||||
|
рані. Потік Са2+ регулюється Са2+-АТФазою |
||||||||||||
|
1. Інозитол 1,4,5-трифосфат зв’язується з ре- |
||||||||||||
|
цептором на ендоплазматичному ретикулумі, що |
мембрани клітини, яка за рахунок енергії АТФ |
|||||||||||
|
призводить до швидкого виходу іонів кальцію з |
відкачує Са2+ з цитоплазми у зовнішнє середо- |
|||||||||||
|
внутрішньоклітинних запасів. Збільшення кон- |
вище (при обміні на іони Na+). Всередині кліти- |
|||||||||||
|
центрації іонів кальцію робить можливим утво- |
ни іони Са2+ депонуються в матриксі міто- |
|||||||||||
|
рення комплексу Са-кальмодулін із чотирма іона- |
хондрій, а в м’язовій тканині — у саркоплазма- |
|||||||||||
|
ми Са2+, який активує молекули білків. Інозитол- |
тичному ретикулумі. Кальцій надходить із зов- |
|||||||||||
|
1,4,5-трифосфат при цьому швидко дефосфори- |
нішнього середовища або внутрішньоклітинних |
|||||||||||
|
лується до інозитол-1,4-дисфосфату та інозитол- |
депо під дією різних стимулів, взаємодіє з Са2+- |
|||||||||||
|
1-фосфату, останні неактивні як вторинні месен- |
зв’язуючим білком цитоплазми кальмодуліном. |
|||||||||||
|
джери (рис. 14.6). |
|
|
|
|
|
|
|
|
|
Одним із внутрішньоклітинних месенджерів |
||
|
|
|
|
|
|
|
|
|
|
гормонального сигналу є іони кальцію, які, крім |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
модифікації ферментативної активності, беруть |
|
|
|
Гормон |
Активований |
участь у виконанні деяких фізіологічних функцій |
||||||||
|
|
|
|
|
|
|
|
|
|
|
рецептор |
клітини (ріст, поділ, згортання крові, передача |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
нервового імпульсу, контрактильна функція). |
|
|
|
|
|
|
|
|
|
|
|
|
|
Передача гормонального сигналу і включення |
|
|
|
|
|
|
|
|
|
|
|
|
|
ефекторних систем відбувається завдяки створен- |
|
|
|
|
|
|
|
|
|
|
|
|
|
ню іонних потоків через плазматичну і внутріш- |
|
|
|
|
|
Фосфоліпаза С |
ньоклітинні мембрани. |
|||||||
|
Фосфоліпіди |
|
|
|
|
|
|
|
|
|
|
Одним із шляхів нагромадження іонів кальцію |
|
|
|
|
|
|
|
|
|
|
|
|
в клітині є відкриття кальцієвих каналів на плаз- |
||
|
мембрани клітини |
|
|
|
|
|
|
|
|
|
|||
|
Вторинні |
|
|
|
|
|
|
|
|
|
|
|
матичних мембранах за допомогою іонотропних |
|
|
|
|
|
|
|
|
|
|
Діацил- |
рецепторів, на які діють медіатори нервової сис- |
||
|
|
|
Інозитол 1,4,5- |
|
|||||||||
|
месенджери |
|
|
|
|
теми. |
|||||||
|
|
|
|
|
|||||||||
|
|
|
|
|
трифосфат |
|
гліцерол |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
Другим шляхом нагромадження кальцію в |
|
|
|
|
|
|
|
|
|
|
|
|
|
цитоплазмі клітин є мобілізація кальцію із внут- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ендоплазматичний |
+ |
|
Са2+ |
+ |
|
рішньоклітинних структур (мітохондрій, ендо- |
||||||
|
|
|
|
|
|
|
плазматичного ретикулума) за допомогою фос- |
||||||
|
ретикулум Са2+ |
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
Протеїнкіназа С |
фатидилінозитолової системи. |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
У результаті підвищується функціональна ак- |
|
|
|
|
|
|
|
Фосфорильовані білки |
||||||
|
|
|
|
|
|
|
тивність і модифікується спрямованість метабо- |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
лізму в клітині. При цьому концентрація іонів |
|
|
|
|
|
|
|
Внутрішньоклітинні ефекти |
||||||
|
|
|
|
|
|
кальцію в ній підвищується в сотні разів. Для по- |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
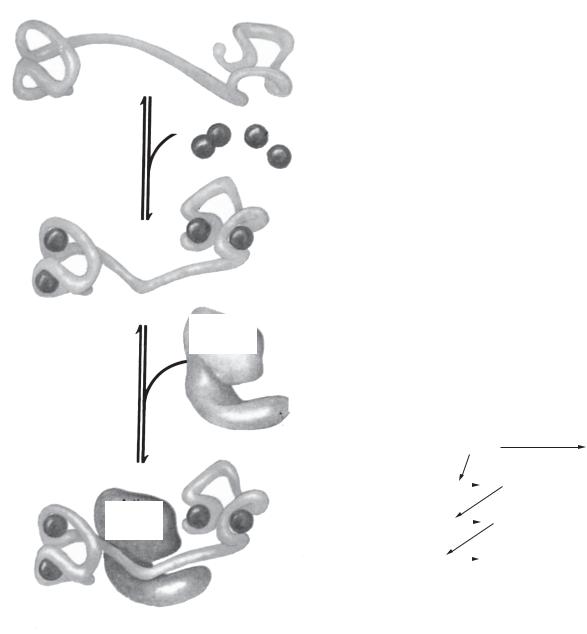
Рис. 14.6. Кальцій/фосфатидилінозитолова система |
вернення клітини у вихідний стан необхідно «пе- |
|||||||||||
|
рекачати» надлишок іонів кальцію всередину |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
ендоплазматичного ретикулума або за межі |
|
2. 1,2-діацилгліцерол активує протеїнкіназу С |
клітини проти градієнта концентрації. Відбу- |
|||||||||||
|
вається цей енергозалежний процес за участю |
||||||||||||
|
— фермент, який фосфорилує білки. Протеїнкіназі |
Са-АТФаз. Установлена структура Са-АТФази |
|||||||||||
|
С необхідні іони кальцію для максимальної ак- |
саркоплазматичного ретикулума, що має 3 фраг- |
|||||||||||
|
тивності, а також фосфоліпід — фосфатидилсе- |
менти. Перший фрагмент, володіючи АТФазною |
|||||||||||
|
рин. |
|
|
|
|
|
|
|
|
|
активністю, гідролізує АТФ і забезпечує енергією |
||
|
3. Синергізм між месенджерами. Два вторин- |
перенос іонів кальцію. Другий фрагмент прони- |
|||||||||||
|
них месенджери — 1,2-діацилгліцерол й інозитол- |
зує товщину мембрани саркоплазматичного ре- |
|||||||||||
|
1,4,5-трифосфат — діють в одному напрямку, по- |
тикулума і формує «канал» для транспорту каль- |
|||||||||||
|
силюючи фосфорилування білків. Приклади |
цію. Третій фрагмент розташований з внутріш- |
|||||||||||
|
клітинних функцій, які залежать від функціону- |
ньої сторони мембрани і виконує роль «заглуш- |
|||||||||||
|
вання фосфо-інозитолового каскаду: |
ки», перешкоджаючи зворотному потоку іонів. |
|||||||||||
|
1. Глікогеноліз у гепатоцитах. |
|
|
|
У цитоплазмі клітин-мішеней існує білок каль- |
||||||||
|
2. Скорочення м’язів. |
|
|
|
модулін, який при підвищенні концентрації іонів |
||||||||
|
3. Агрегація тромбоцитів і вивільнення ними |
Кальцію зв’язується з 4 іонами Кальцію, змінює |
|||||||||||
|
серотоніну. |
|
|
|
|
|
|
|
|
|
конформацію і набуває здатності активувати |
||
199
Кальмодулін
4Са2+
Кальмодулін + 4Са2+
Неактивний
фермент (кіназа)
Активний
фермент (кіназа)
в) активацією фосфоліпази С із залученням фосфоінозитидної системи і мобілізацією внутрішньоклітинного Са;
г) надходженням Са із позаклітинного середовища через іонні канали.
Трансформація гормонального сигналу в біохімічні реакції клітин відбувається через протеїнкінази: цАМФ-залежні; цГМФ-залежні; Са/кальмодулінзалежні; Са/фосфоліпідзалежні; тирозинзалежні протеїнкінази.
цАМФ
цАМФ активує протеїнкінази, які існують у двох формах (активній і неактивній). Неактивна форма складається з 2 субодиниць (каталітичної та рецепторної). R2C2-комплекс не виявляє ферментативної активності. цАМФ з’єднується з рецепторною субодиницею і відокремлює її від каталітичної субодиниці:
4 цАМФ + R2C2 → R2 (4 цАМФ) +2C,
де C — каталітична субодиниця; R — регуляторна субодиниця.
Протеїнкінази каталізують фосфорилування відповідних ферментів, переводячи тим самим їх у неактивний чи активний стан.
аденілатциклаза фосфодіестераза
АТФ цАМФ АМФ
цАМФ АМФ
Протеїнкіназа |
|
Протеїнкіназа |
||||
(неакт.) |
|
|
|
|
(акт.) |
|
Кіназа фосфори- |
|
Кіназа фосфори- |
||||
лази (неакт.) |
|
|
|
лази (акт.) |
||
|
|
|
||||
Фосфорилаза |
|
Фосфорилаза |
||||
(неакт.) |
|
|
|
(акт.) |
||
Рис. 14.7. Комплекс 4Са2+-кальмодулін
групу кальмодулінзалежних протеїнкіназ, які фосфорилують ряд ферментних білків, модифікуючи їхню активність (рис. 14.7).
Вторинні месенджери
Найбільш розповсюдженими вторинними месенджерами є цАМФ та фосфатидилінозитол. цАМФ є посередником у передачі гормонального сигналу в клітині, він утворюється з АТФ під дією активної форми аденілатциклази. Сигнал від рецептора гормону на аденілатциклазу переноситься через G-білок, вбудований у мембрани, що має ГТФазну активність. Комплекс G-білок із ГТФ активує аденілатциклазу, а комплекс G-білок із ГДФ не має такої дії. Збільшення внутрішньоклітинної концентрації вторинних месенджерів досягається:
а) активацією аденілатциклази; б) активацією гуанілатциклази;
цГМФ
цГМФ функціонує як «вторинний месенджер» у кишечнику, серці, мозку, судинах і нирках. У кишечнику і нирках його дія приводить до зміни транспорту іонів й збереження води. У гладкому та серцевому м’язі цГМФ викликає розслаблення (релаксацію), у мозковій тканині може брати участь у її розвитку, а для зрілого мозку — у функціонуванні. Принаймні два ізоферменти гуанілатциклази продукують цГМФ із ГТФ. Ця реакція аналогічна реакції, яка каталізується аденілатциклазою.
гуанілатциклаза
ГТФ —————→ цГТФ + H4P2O7
Один з ізоферментів — інтегральний білок плазматичної мембрани із доменом рецептора на зовнішній поверхні мембрани і доменом цГМФ на цитозольній стороні — має три ділянки: рецепторну, внутрішньомембранну, каталітичну. Цей ізофермент гуанілатциклази активується зв’язуванням із гормоном — передсердним натрійуретичним фактором, який виділяється клітинами передсердя серця, коли збільшується на-
200
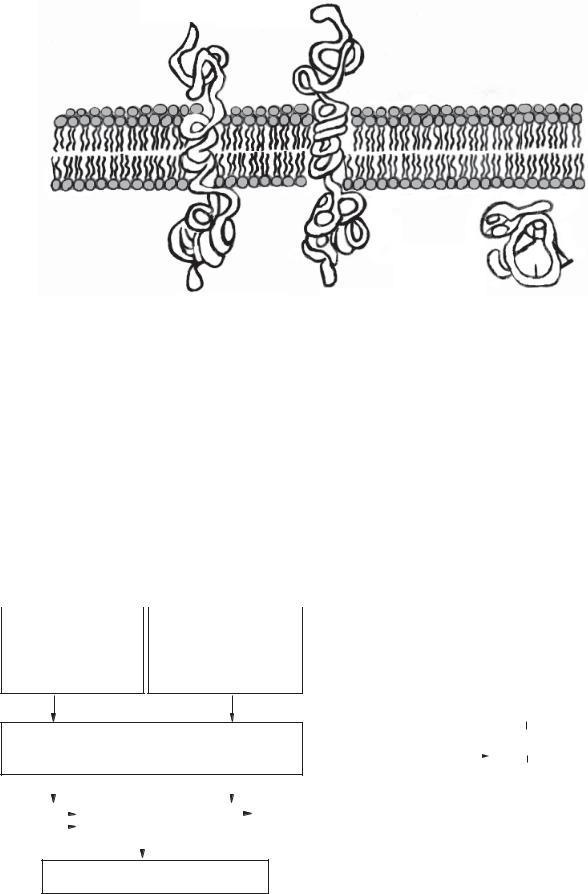
Рецептор передсердного |
Рецептор |
натрійуретичного фактора |
ендотоксину |
Позаклітинні лігандзв’я- зуючі домени рецептора
Внутрішньоклітинні каталітичні домени (утворені циклічним ГМФ)
|
|
|
|
|
|
|
Гем |
|
|
|
|
|
|
COOH |
Розчинна гуанілатциклаза |
|
|
|
|
|
|
||
|
|
|
|
|
|
||
|
|
COOH |
|||||
|
|
|
|
|
|||
Рис. 14.8. Два типи гуанілатциклаз (ізозимів), що беруть участь у сигнальній трансдукції |
|||||||
повнення передсердя кров’ю (blood volume |
ним ендотоксином (невеликий пептид), що про- |
||||||
stretches the atrium). Принесений кров’ю перед- |
дукується Е. coli. У результаті збільшення кон- |
||||||
сердний натрійуретичний фактор активує гу- |
центрації цГМФ призводить до зменшення реаб- |
||||||
анілатциклазу в клітинах нирок. Підвищення |
сорбції води епітелієм кишечнику, ця дія токсину |
||||||
цГМФ збільшує виділення нирками Na+ і, |
спричинює діарею (рис. 14.8). |
||||||
відповідно, води. Втрата води знижує кров’я- |
|
Другий і явно відмінний ізофермент гуанілат- |
|||||
ний тиск. Гладкі м’язи судин також мають ре- |
циклази — цитозольний білок, який щільно асо- |
||||||
цептор натрійуретичного фактора — гуанілат- |
ціюється з гемом (розчинний білок). Цей фермент |
||||||
циклазу. У результаті зв’язування з рецептором |
активується природною сполукою — оксидом |
||||||
натрійуретичного фактора відбувається розслаб- |
азоту NO (II) і судинорозширювальними нітро- |
||||||
лення судин, через що тиск крові знижується. |
похідними сполуками, такими як нітрогліцерин |
||||||
Подібний рецептор гуанілатциклази в плаз- |
і нітропрусид (використовуються в лікуванні хво- |
||||||
матичній мембрані епітеліальних клітин кишеч- |
роб серця). |
|
|||||
нику активується термостабільним бактеріаль- |
|
Судинорозширювальні нітропохідні спонтан- |
|||||
|
|
|
|
|
но розщеплюються, утворюючи NO (рис. 14.9). |
||
|
|
|
|
|
|
В організмі людини NO (II) утворюється з ар- |
|
|
|
Негативні модулятори: |
|
||||
|
|
гініну за допомогою Са2+-залежної оксидази зі |
|||||
Позитивні модулятори: |
|
гемопротеїни, |
|||||
|
змішаними функціями — NO-синтази, яка при- |
||||||
|
аналоги аргініну, |
||||||
аргінін, ацетилхолін, |
|
сутня у багатьох тканинах: |
|||||
|
супероксидрадикали, |
||||||
брадикінін |
|
етанол, глюкокортикоїди, |
|
|
|
||
|
|
|
|
|
|||
|
|
|
НПЗС |
|
|
|
|
+ О2 + НАДФН+Н+
Аргінін  Цитрулін + NO NO-синтаза
Цитрулін + NO NO-синтаза
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
NO + е |
|
NO– |
|
|
NO + О2– |
|
|
ОNOО– |
|
|
|
|
|
|
|||||
|
|
|
|||||||
NO – е |
|
|
NO+ |
|
|
(пероксинітрит) |
|||
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
Інактивація — утворення NО2– і NО3–, які не є вазодилататорами
Рис. 14.9. Утворення і розпад оксиду азоту
|
|
O |
|
|
|
O |
|
|
|
= |
|
|
|
|
|
H2N–CH–C–OH |
= |
|
|||||
|
CH2 |
|
|
H2N–CH–C–OH |
|
||
|
|
|
|||||
|
|
|
|
CH2 |
|
||
|
CH2 |
+ O2 |
|
|
CH2 |
+ NO |
|
|
|
|
|||||
|
|
|
|
|
|||
CH |
2 |
|
|
CH2 |
|||
|
|
|
|
|
|
|
|
N H |
|
|
|
|
|
||
|
|
N H |
|
||||
|
C=NH |
|
|
|
|||
|
|
|
|
|
|
||
|
|
|
C=O |
|
|||
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
NH2 |
|
|
|
|
|
||
|
|
NH2 |
|
||||
Аргінін |
|
|
|
||||
|
|
Цитрулін |
|
||||
NO дифундує з цих клітин у сусідні, де зв’я- зується з гемом гуанілатциклази й активує фермент, який продукує цГМФ. У серці цГМФ знижує силу серцевих скорочень за рахунок стимулювання іонних насосів, які забезпечують низь-
201
ку концентрацію Са2+ у цитозолі. Це розслаблення серцевого м’яза аналогічне дії нітрогліцерину, який використовується для полегшення стенокардії, болю, спричиненого скороченням серця, позбавленого О2 через блокування коронарних артерій.
Участь оксиду азоту в функціях органів і тканин
1.Посилення релаксації та діастолічного розтягнення шлуночків, підвищення хронотропної та ізотропної відповіді.
2.Релаксація стравоходу, шлунка, дванадцятипалої кишки, посилення перистальтики кишечнику, надходження жовчі, захист слизової шлунка.
3.Синтез NO в ендотелії легеневих судин, епітелії верхніх дихальних шляхів, бронходилатація.
4.Регуляція ниркової гемодинаміки і гломерулярної фільтрації, інгібування транспорту Nа і збільшення його екскреції.
5.Зниження адгезії та агрегації тромбоцитів, лейкоцитів, факторів росту, антимітогенна й антипроліферативна дія.
Оксид азоту (NO) — нестабільна сполука короткочасної дії — протягом кількох секунд після утворення; потім NO піддається окисненню до нітрату або нітриту. Оскільки перетворення нітрогліцерину в NО відбувається повільно, то нітрогліцерин спричинює тривале (пролонговане) розслаблення серцевого м’яза.
Вважають, що більшість ефектів цГМФ пов’я- зана з цГМФ-залежною протеїнкіназою — так званою протеїнкіназою G.
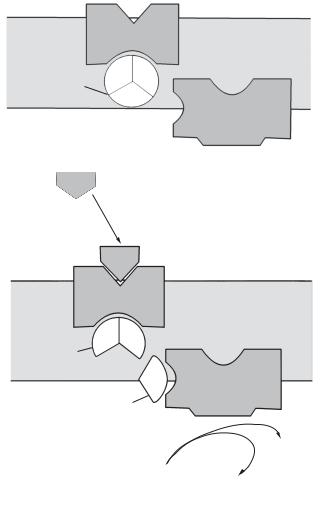
G-білок
Яке значення G-білка в клітинній активності? Перетворення ГТФ на ГДФ відбувається під дією ферменту — ГТФази, яка гідролізує ГТФ. Скорочено «білок, зв’язуючий гуанілові нуклеотиди» стали називати просто G-білок. Він складається з трьох субодиниць: α , β , γ . У стані спокою ці субодиниці утворюють комплекс, а ГДФ пов’язаний з α -субодиницею. Після того, як первинний месенджер (гормон чи медіатор) приєднується до рецептора, конформація останнього змінюється, він, у свою чергу, зв’язує G-білок. У результаті цієї взаємодії α -субодиниця вивільняє ГДФ. Тепер ГТФ, концентрація якого в клітинах вища, змінює форму α -субодиниці й активує її. Активована і зв’язана з ГТФ α -субодиниця відокремлюється від β - і γ -субодиниць і шляхом дифузії переміщується по внутрішній поверхні плазматичної мембрани, поки не зв’яжеться з ефектором (аденілатциклазою). Зазвичай через кілька секунд α -субодиниця гідролізує ГТФ до ГДФ і тим самим виключається (рис. 14.10, табл. 14.4).
|
Рецептор |
|
||
G-білок |
γ |
|
β |
|
|
α |
|
||
|
|
Аденілатцик- |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
лаза неактивна |
|
ГДФ |
|
||
Рецептор
γβ
G-білок
αАденілатциклаза активна
ГТФ
цАМФ
АТФ
+РРі
Рис. 14.10. G-Комплекс цАМФ аденілатциклаза
Таким чином, ефекторні системи клітин представлені величезною кількістю протеїнкіназ, які модифікують метаболізм у клітинах шляхом АТФ-залежного фосфорилування ферментних білків. Існує кілька груп протеїнкіназ:
1.цАМФ-залежні протеїнкінази. До гормонів, що активують аденілатциклазу (діють
через GS-білок), належать адреналін, глюкагон, вазопресин і практично всі тропні гормони гіпофіза. До гормонів, що інгібують адені-
латциклазу (діють через GI-білок), належать ангіотензин, ацетилхолін, опіоїдні пептиди, соматостатин.
2.цГМФ-залежні протеїнкінази.
3.Са2+/кальмодулінзалежні протеїнкінази.
4.Са2+/фосфоліпідзалежні протеїнкінази.
5.Тирозинзалежні протеїнкінази.
Перші 4 групи протеїнкіназ фосфорилують залишки серину або треоніну, що входять у структуру ферментних білків, а тирозинзалежні протеїнкінази фосфорилують залишки тирозину. Характерною рисою дії тирозинзалежних протеїнкіназ є те, що вони не потребують вторинних посередників, а самі зв’язуються з гормоном на зовнішній поверхні мембран, пронизують товщу мембрани і фосфорилують ферментні білки за
202
|
|
|
|
|
Таблиця 14.4 |
|
Приклади фізіологічних ефекторів за участю G-білків |
|
|||
|
|
|
|
|
|
Гормон |
Тип клітин-мішеней |
G-білок |
Ефектор |
|
Ефект |
|
|
|
|
|
|
Адреналін, глюкагон |
Клітини печінки |
Gs |
Аденілатциклаза |
|
Розщеплення глікогену |
Адреналін, глюкагон |
Жирова клітина |
Gs |
Аденілатциклаза |
|
Розщеплення жирів |
Лютропін |
Клітини яєчників |
Gs |
Аденілатциклаза |
|
Посилення синтезу |
|
|
|
|
|
естрогенів і прогестерону |
Вазопресин |
Клітини нирок |
Gs |
Аденілатциклаза |
|
Затримка води нирками |
Ангіотензин |
Гладкі м’язи судин |
Gq |
Фосфоліпаза С |
|
М’язові скорочення, |
|
|
|
|
|
підвищення кров’яного |
|
|
|
|
|
тиску |
Ацетилхолін |
Клітини серцевого |
Gi |
Калієві канали |
|
Уповільнення |
|
м’яза |
|
|
|
і ослаблення |
|
|
|
|
|
серцевих скорочень |
Енкефаліни, |
Нейрони головного |
Gi/G0 |
Са- і К-канали, |
|
М’язові скорочення, |
ендорфіни, |
мозку |
|
аденілатциклаза |
|
підвищення кров’яного |
опіоїди |
|
|
|
|
тиску |
|
|
|
|
|
|
Примітки. Gs-білки — стимулюючі, які активують аденілатциклазу; Gi-білки — інгібуючі, які інгібують аденілатциклазу; Gq-білки, які активують фосфоліпазу С.
участю фрагмента, що виходить у цитоплазму |
ми ділянками ДНК, у результаті чого активу- |
|
клітин. Типовим лігандом для цієї групи протеїн- |
ються специфічні гени і транскрипція матричних |
|
кіназ є інсулін. |
РНК, які на рибосомах забезпечують синтез пев- |
|
|
них ферментних білків, що реалізують гормо- |
|
|
нальний ефект шляхом модифікації метаболізму. |
|
|
Оскільки в реалізацію ефекту гормонів цитоплаз- |
|
|
матичного механізму дії залучається біосинтез |
|
14.2. МОЛЕКУЛЯРНО-КЛІТИННІ |
білка, то прояв цього ефекту за часом тривалі- |
|
ший, ніж у гормонів, що зв’язуються з метабо- |
||
МЕХАНІЗМИ ДІЇ СТЕРОЇДНИХ |
||
тропними рецепторами, і значно поступається |
||
І ТИРЕОЇДНИХ ГОРМОНІВ |
гормонам, що зв’язуються з іонотропними рецеп- |
|
|
торами. Більшість гормонів діють за мембранним |
|
На відміну від гормонів білково-пептидної |
механізмом. Лише стероїдні гормони й тироксин |
|
мають яскраво виражений цитоплазматичний |
||
природи і похідних амінокислот, рецептори яких |
механізм дії (див. рис. 14.4). |
|
розташовані на мембранах клітин-мішеней, ре- |
|
|
цептори стероїдних і тиреоїдних гормонів локалі- |
|
|
зовані в цитоплазмі і ядрі клітин. Внаслідок своєї |
|
|
ліпофільності, ці гормони проникають через плаз- |
|
|
матичну мембрану клітини в цитоплазму і зв’я- |
|
|
зуються зі специфічними рецепторами, утворюю- |
КОНТРОЛЬНІ ПИТАННЯ |
|
чи гормон-рецепторні комплекси, які транспорту- |
|
|
ють гормони до певних субклітинних структур. |
|
|
Гормони цитоплазматичного механізму дії впли- |
1. Гормони: загальна характеристика; роль |
|
вають, головним чином, на генетичний апарат |
гормонів та інших біорегуляторів у системі |
|
клітини, тобто цей комплекс транспортується в |
міжклітинної інтеграції функцій організму люди- |
|
ядро клітини, де гормон зв’язується з певними |
ни. |
|
сайтами ДНК і підсилює або гальмує синтез пев- |
2. Класифікація гормонів і біорегуляторів: |
|
них мРНК, що забезпечують синтез ферментних |
||
відповідність структури та механізмів дії гор- |
||
білків de novo. Цитоплазматичні рецептори скла- |
||
даються з: а) домену, що з’єднується з гормоном; |
монів. |
|
б) домену, що з’єднується з певними ділянками |
3. Реакція клітин-мішеней на дію гормонів. |
|
ДНК; в) домену, що з’єднується з додатковими |
Мембранні (іонотропні, метаботропні) та цито- |
|
регуляторами транскрипції; г) Zn-вмісних до- |
зольні рецептори. |
|
менів — «цинкових пальців», які виступають як |
4. Біохімічні системи внутрішньоклітинної пе- |
|
регулятори транскрипції й мають вигляд випи- |
||
редачі гормональних сигналів: G-білки, вторинні |
||
нань ділянок поліпептидного ланцюга рецептор- |
||
ного білка, сформованих за рахунок зв’язуван- |
посередники (цАМФ, Са2+/кальмодулін, ІФ3, |
|
ДАГ). |
||
ня іонів Цинку з амінокислотами. |
|
|
Активований гормон-рецепторний комплекс |
5. Молекулярно-клітинні механізми дії стероїд- |
|
переміщається в ядро, зв’язується зі специфічни- |
них і тиреоїдних гормонів. |
203

Глава 15. БІОХІМІЯ ГОРМОНАЛЬНОЇ РЕГУЛЯЦІЇ
15.1. ГОРМОНАЛЬНА РЕГУЛЯЦІЯ МЕТАБОЛІЗМУ ТА КЛІТИННИХ ФУНКЦІЙ: ГОРМОНИ ГІПОТАЛАМОГІПОФІЗАРНОЇ СИСТЕМИ, ПІДШЛУНКОВОЇ І ЩИТОПОДІБНОЇ ЗАЛОЗ
В ієрархії гормональних систем гіпоталамус спричинює регуляторний вплив на функцію гіпофіза за рахунок системи нейропептидів (ліберинів і статинів). Важливу роль у продукції гормонів гіпофіза і нейропептидів відіграє проопіомеланокортин — білок із молекулярною масою 29 000 Да, який є попередником багатьох гормонів. Відбувається це шляхом протеолізу під впливом специфічних протеаз, які гідролізують пептидні зв’язки молекули проопіомеланокортину між певними залишками амінокислот, внаслідок чого утворюються ті чи інші гормони гіпофіза. Родина пептидів проопіомеланокортину (ПОМК) складається з пептидів, які діють або як гормони (кортикотропін, ліпотропін, меланотропін), або як нейромедіатори чи нейромодулятори; ПОМК синтезує приблизно 5 % клітин передньої частки гіпофіза і всі клітини проміжної частки. Регуляція синтезу і секреції ПОМК у цих відділах різна. Різні пептидні гормони можуть бути виділені з того самого попередника за допомогою протеолітичного процесингу. Сегменти прогормону відщеплюються і піддаються посттрансляційній модифікації шляхом глікозування, ацетилювання і фосфорилування. Подальше розщеплення ПОМК у передній і проміжній частках гіпофіза відбувається на ділянці між АКТГ (адренокортикотропний гормон) і β -ЛТГ (β -ліпотропний гор-
мон) (рис. 15.1).
Механізм дії та функції гормонів гіпофіза
За хімічною будовою тиреотропін, фолі-
тропін, лютропін, гонадотропін людини (пла-
цента) є глікопротеїнами. Вони складаються з двох субодиниць: α і β . α -Cубодиниці в них однакові, а β -субодиниці розрізняються, визначаючи специфічність гормонів. Інші гормони (кортикотропін, пролактин, гормон росту) є простими білками, що мають один поліпептидний ланцюг, а вазопресин і окситоцин — це циклічні октапептиди.
Усі тропні гормони впливають на функції периферичних залоз або безпосередньо на периферичні тканини після зв’язування з їх мембранними рецепторами й активації аденілатциклази або гуанілатциклази. Саме тому гормони гіпофіза називаються тропними гормонами (від грецьк. tropos — поворот, напрямок).
Передня частка гіпофіза
Перебуваючи під контролем гормонів гіпоталамуса, вона секретує ряд тропних гормонів.
Кортиколіберин є основним фактором, який регулює вивільнення ПОМК із передньої частки гіпофіза. Він діє через цАМФ-систему, яка вимагає присутності іонів Са2+. Глюкокортикоїди інгібують утворення кортиколіберину та його секрецію.
Мінорні ефекти на секрецію ПОМК передньої частки справляють вазопресин і α -адренергічні агенти, а також через ЦНС — серотонін, ацетихолін і γ -аміномасляна кислота (ГАМК, γ -аміно- бутират).
Тиреотропін (тиреотропний гормон) — глікопротеїн. Контролює розвиток і функцію щитоподібної залози.
26 |
|
|
|
48 |
|
|
|
12 |
|
|
|
|
|
40 |
|
|
|
14 |
|
|
|
|
21 |
|
|
|
|
|
40 |
|
|
|
18 |
|
|
|
|
31 |
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Проопіомеланокортин |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
40–134 |
|
|
|
|
|
40–134 |
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
γ -МСГ |
|
|
|
|
|
|
Кортикотропін |
|
|
|
|
β -Ліпотропін |
|
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α -МСГ |
|
|
КППД |
|
|
|
|
γ -ЛТГ |
|
|
β -Ендорфіни |
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1–13 |
|
|
18–39 |
|
|
|
42–101 |
|
|
|
104–134 |
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β -МСГ |
|
|
γ -Ендорфіни |
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
104–118 |
|
||
Рис. 15.1. Продукти розщеплення проопіомеланокортину: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||
МСГ — меланоцитстимулювальний гормон; |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α -Ендорфіни |
|
|||||||||||||||||||||||
КПП — кортикотропіноподібний пептид; |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||
ЛПГ — ліпотропін |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
104–117 |
|
|||||||||
204
1. Сприяє нагромадженню йоду в щито- |
|
Кортиколіберин (гіпоталамус) |
|||
подібній залозі. |
|
|
|
+ |
|
|
|
|
|||
2. Прискорює включення йоду в молекулу ти- |
|
|
|
||
розину на білковій «подушці» — йодтиреогло- |
– |
|
|
|
|
|
Кортикотропін (гіпофіз) |
||||
буліні. |
|
||||
|
|
|
|
||
3. Активує протеолітичні ферменти, які вивіль- |
|
– |
|
+ |
|
няють тиреоїдні гормони з йодтиреоглобуліну. |
|
|
|
||
|
|
|
|
||
4. Впливає на енергетичний обмін, підсилює |
|
|
Кортикостероїди |
||
нагромадження оксидоредуктаз із SH-групами |
Рис. 15.2. Регуляція синтезу і секреції кортикотропіну |
||||
(адаптація, посилення вільного окиснення). |
|||||
Симптоматика гіперпродукції тиреотропіну по- |
за принципом негативного зворотного зв’язку |
||||
дібна дії гормонів щитоподібної залози (дифузій- |
Примітка. + — активація; – — інгібування. |
||||
|
|
|
|
||
ний токсичний зоб). Гіпопродукція тиреотропіну |
|
|
|
|
|
спричинює розвиток тиреотропного (гіпофізар- |
При гіперпродукції кортикотропіну збільшу- |
||||
ного) карликового росту. Рівень холестеролу при |
єтьсяутворенняглюкокортикоїдів, головнимчином |
||||
гіпофізарній мікседемі в нормі чи знижений, при |
кортизолу. Розвивається хвороба Іценко — Ку- |
||||
мікседемі щитоподібної залози — гіперхолесте- |
шинга (гіперкортицизм). Гіпопродукція кортикот- |
||||
ролемія. |
ропіну пов’язана з пухлиною, інфекцією чи інфар- |
||||
Фолітропін (фолікулостимулювальний гор- |
ктом гіпофіза, спричинює протилежні зрушення. |
||||
мон) — глікопротеїн зв’язується зі специфічними |
Соматотропін (гормон росту) — поліпептид. |
||||
рецепторами на плазматичних мембранах клі- |
Секреція має пульсуючий характер. Один із най- |
||||
тин-мішеней — фолікулярних клітин яєчників і |
більших піків продукції відбувається одразу ж |
||||
клітин Сертолі в сім’яниках. |
після засинання. |
||||
1. Сприяє дозріванню фолікулів у яєчниках у |
1. Спричинює ріст скелета за рахунок труб- |
||||
самок, підсилює секрецію естрогенів. |
частих кісток, затримує їхнє скостеніння. Ріст |
||||
2. Сприяє сперматогенезу в самців за рахунок |
здійснюється в результаті регуляції обміну білка, |
||||
синтезу андрогензв’язувального білка, який бере |
кальцію, |
фосфору. |
|||
участь у транспорті тестостерону, необхідного |
2. Викликає формування особливостей будо- |
||||
для сперматогенезу. |
ви тіла, збільшення ваги за рахунок активації |
||||
Лютропін (лютеїнізуючий гормон) — гліко- |
біосинтезу білка, нуклеїнових кислот. |
||||
протеїн. |
3. Впливає на вуглеводний обмін, сприяє за- |
||||
1. У самок — зрілий фолікул, забезпечує до- |
своєнню вуглеводів, стимулюючи секрецію інсу- |
||||
зрівання, овуляцію й утворення жовтого тіла, ут- |
ліну. Підсилює розщеплення вуглеводів, затри- |
||||
ворення прогестерону клітинами жовтого тіла. |
мує їхнє перетворення на ліпіди (жири). |
||||
2. У самців — клітини Лейдига, де гормон сти- |
|||||
мулює утворення тестостерону, сім’яники, де гор- |
4. Сприяє мобілізації жиру з депо й окиснен- |
||||
ню його в тканинах. |
|||||
мон стимулює ріст інтерстиціальних клітин. |
Соматотропін чинить прямий ефект на кістко- |
||||
Пролактин (лактотропін, лактогенний гор- |
|||||
ву тканину, острівці Лангерганса. На жирову |
|||||
мон) — білок. |
тканину, м’язову тканину, хрящі гормон впли- |
||||
1. Стимулює розвиток молочних залоз і лак- |
|||||
ває через особливий білковий фактор — сомато- |
|||||
тацію. |
медин, який синтезується в печінці. Гіперфункція |
||||
2. Стимулює ріст внутрішніх органів. |
|||||
соматотропіну у дитячому віці викликає посиле- |
|||||
3. Гальмує ефект лютеїнізуючого гормону — |
|||||
ний ріст — гігантизм, у дорослих — розвиток ак- |
|||||
овуляцію (зберігає жовте тіло). |
ромегалії (від грецьк. аcros — крайній, кінцівка, |
||||
4. У жировій тканині активує ліпогенез. |
|||||
megas — великий), характеризується непропор- |
|||||
Кортикотропін (адренокортикотропний гор- |
|||||
ційним інтенсивним ростом окремих частин тіла. |
|||||
мон, поліпептид), орган-мішень — надниркові |
Гіпофункція соматотропіну спричинює розви- |
||||
залози і жирова тканина. Необхідний для фор- |
|||||
ток гіпофізарного нанізму (карликовий зріст із |
|||||
мування структури і нормального функціону- |
порушенням розвитку всіх органів і тканин). |
||||
вання кори надниркових залоз, а також синте- |
|
|
Проміжна частка |
||
зу і секреції кортикостероїдів. |
|
|
|||
1. Підсилює проникнення глюкози в тканину |
Вона містить гормонально активні речовини |
||||
надниркових залоз, що перетворюється на аце- |
— похідні пропіомеланокортину. Ця частка |
||||
тил-КоА, а останній — на холестерол і кортико- |
бідна на кровоносні судини, гіпоталамо-гіпофі- |
||||
стероїди. |
зарна портальна система її не досягає, і тому на |
||||
2. Активує пентозофосфатний шлях, джерело |
неї не впливає кортиколіберин. У проміжній |
||||
НАДФН + Н+. |
частці немає рецепторів глюкокортикоїдів, тому |
||||
3.Активує ліполіз у жировій тканині. останні не регулюють продукцію ПОМК. Про-
4.Підвищує рівень холестеролу в корі надміжна частка гіпофіза іннервована дофамінергіч-
ниркових залоз.
5. Синтез і секреція кортикотропіну регулюється за принципом негативного зворотного зв’язку за допомогою кортикостероїдів (рис. 15.2):
ними волокнами, а також містить серотонінергічні, катехоламінергічні нервові закінчення. Агоністи дофаміну знижують, а антагоністи — підвищують кількість мРНК ПОМК і секрецію пептидів ПОМК. Вивільнення ПОМК у про-
205
міжній частці стимулюється серотоніном і β -адре- |
2. Вазопресин активує гіалуронідазу, яка ка- |
|
нергічними агентами. |
талізує розщеплення гіалуронової кислоти, через |
|
α -Ліпотропін (ліпотропний гормон, |
що збільшується проникність клітинних мемб- |
|
ран, отже, прискорюється реабсорбція H2O з пер- |
||
91 амінокислота) |
||
Ліпотропін активує ліполіз в адипоцитах жи- |
винної сечі й тим самим зменшується діурез, сеча |
|
стає більш концентрованою. |
||
рової тканини і мобілізацію жирних кислот |
При гіперпродукції вазопресину розвивають- |
|
(цАМФ-посередник). Вважають, що фізіологічна |
||
ся набряки (олігоурія, синдром Пархоша), при |
||
дія його незначна. Ліпотропін бере участь в ут- |
гіпопродукції — нецукровий діабет (поліурія до |
|
воренні α -, β -, γ -, і δ -ендорфінів, які виконують |
10 л на добу, супроводжується спрагою — полі- |
|
роль нейромедіаторів, ендогенних знеболюваль- |
дипсія). |
|
них факторів і модуляторів важливих психо- |
Окситоцин (пептид) |
|
фізичних процесів у пептидергічних структурах |
||
1. Сприяє проникненню іонів Ca2+ у клітини. |
||
головного мозку. Ендорфіни зв’язуються в голов- |
Нагромадження іонів Ca2+ підсилює процеси ско- |
|
ному мозку з тими самими рецепторами цент- |
рочення (матка, бронхи, стінки кишечнику, мо- |
|
ральної нервової системи, що й морфінові опіати, |
лочна залоза). |
|
вони можуть відігравати роль в ендогенній регу- |
2. Блокує активність ацетилхолінестерази, |
|
ляції чутливості до болю, мають вищу актив- |
||
підсилюючи цим збудливість м’язів і силу їх ско- |
||
ність, ніж морфін. |
рочень. |
|
Меланоцитостимулюючі гормони (α -, β -, γ - |
||
МСГ) — вони стимулюють функціональну ак- |
3. Стимулює в ендометрії утворення проста- |
|
гландинів, які активують скорочення гладких |
||
тивність меланоцитів шкіри, збільшують її піг- |
м’язів. |
|
ментацію. Особи з низьким рівнем глюкокорти- |
У фізіологічних дозах використовується для |
|
коїдів (хвороба Аддісона) мають посилену |
||
стимуляції пологової діяльності у жінок. |
||
пігментацію шкіри, пов’язану з підвищенням ак- |
|
|
тивності МСГ у плазмі. |
|
Задня частка гіпофіза (нейрогіпофіз) |
Гормони щитоподібної залози |
|
|
Вона продукує гормони, які регулюють вод- |
У щитоподібній залозі здійснюються синтез і |
ний баланс (вазопресин) і викид молока з лак- |
секреція йодотиронінів — тироксину (Т4) і три- |
туючої молочної залози (окситоцин). |
йодотироніну (Т3). Йодотироніни входять до |
Синтез гормонів здійснюється в гіпоталамусі |
складу білка тиреоглобуліну, який міститься в |
— по аксонах вони переносяться в гіпофіз. Мож- |
колоїді фолікулів щитоподібної залози. Секреція |
ливо, у гіпофізі здійснюється дозрівання гор- |
і синтез перебувають під контролем тиреотропі- |
монів, а не тільки їх зберігання. Суть такого ме- |
ну і перебігають у кілька стадій: |
ханізму полягає, імовірно, у тому, що він дозво- |
1. Утворення з йодиду «активного» йоду з |
ляє обминати гематоенцефалічний бар’єр. |
участю йодопероксидази. Акцептором електронів |
Вазопресин (антидіуретичний гормон, АДГ) |
служить Н2О2. Активний йод здатний йодувати |
— пептид. |
тирозин. |
Контролює водний баланс організму, осмо- |
2. Йодування тирозину в складі тиреоглобу- |
тичний тиск крові. Молекулярний механізм дії |
ліну з участю тирозинйодинази. При цьому ут- |
вазопресину визначається наявністю двох типів |
ворюються монойодтирозин або дийодтирозин. |
рецепторів (табл. 15.1): |
3. Окисна конденсація моно- і дийодтирозинів |
V1 — рецептор, локалізований на мембранах |
з утворенням трийодотироніну і тироксину в мо- |
гепатоцитів, тромбоцитів; |
лекулі тиреоглобуліну. Процес здійснюється на |
V2 — рецептор, локалізований на мембранах епі- |
поверхні тирозинйодинази. Гормони щитоподіб- |
теліальних клітин трубочок і петель Генлі нефронів. |
ної залози зберігаються в складі молекули тирео- |
1. Вазопресин стимулює скорочення гладкої |
глобуліну в колоїдній речовині фолікула. |
мускулатури судин, що спричинює підвищення |
4. Поглинання тиреоглобуліну з колоїду клі- |
кров’яного тиску. |
тинами епітелію і переміщення його до зовнішньої |
|
Таблиця 15.1 |
Спрямованість дії вазопресину
Рецептори |
Локалізація дії |
Механізм дії |
Ефекти |
|
|
|
|
V2 |
Нирки (дистальна частина |
Посилення |
Антидіуретична дія |
|
нефрону) |
реабсорбції Н2О |
|
V2 |
Система |
Підвищення вмісту в крові |
Гемостатична дія |
|
згортання крові |
антигемофільного глобуліну А |
|
V1 |
Тромбоцити |
Стимуляція агрегації |
Стимуляція |
|
|
та дегрануляції тромбоцитів |
тромбоутворення |
V1A |
Артеріальні судини |
Підвищення тонусу |
Вазопресорна дія |
|
|
гладких м’язів судин |
|
V1B |
Передня частка гіпофіза |
Стимуляція вивільнення |
Стимуляція вивільнення |
|
|
кортикотропіну |
гідрокортизону |
|
|
|
|
206
поверхні мембрани, оточеної позаклітинною рідиною.
5. Секреція йодотиронінів здійснюється завдяки гідролізу протеазами тиреоглобуліну, з якого вивільняються в кров тироксин (Т4) і трийодоти-
ронін (Т3).
У крові йодотироніни утворюють комплекси з тироксинзв’язувальним глобуліном (75 % цирку-
люючого гормону), з альбуміном (10 %), з тиреозв’язувальним преальбуміном плазми (15 %), потім транспортуються до периферичних тканин. З білками плазми Т3 зв’язаний приблизно в 3–5 разів слабше, ніж Т4, тому біологічний ефект Т3 приблизно в 3–5 разів сильніший за Т4.
Приблизно 99,96 % циркулюючого тироксину пов’язано з білками. У такому стані гормон не має безпосередньо метаболічної активності, але перебуває в рівновазі з 0,04 % вільного гормону. Саме така форма гормону здатна до активності й бере участь у метаболізмі. Велика кількість Т4 зберігається в колоїдній речовині численних фолікулів щитоподібної залози. Хоча тироксин — основний продукт щитоподібної залози, він не є активною формою гормону. Вища спорідненість ядерних рецепторів до Т3 порівняно з Т4 робить Т3 сильнішим гормоном. При нормальному харчуванні й функціонуванні щитоподібної залози близько 15 % циркулюючого Т3 надходить із залози. Більша частина циркулюючого гормону утворюється в результаті періодичного дейодування Т4. Ферментні системи печінки, нирок та інших тканин контролюють продукцію активного Т3 (залежно від певних умов чи дії лікарських препаратів) або сприяють його переходу в неактивну форму — реверсивний Т3 (рТ3).
I |
|
I |
|
|
HO |
O |
CH2 |
CH |
COOH |
I |
|
I |
NH2 |
|
|
|
|
||
|
Тироксин (Т4) |
|
|
|
I |
|
I |
|
|
HO |
O |
CH2 |
CH |
COOH |
|
|
I |
NH2 |
|
|
|
|
|
|
Трийодотиронін (Т3) активний |
||||
I |
|
I |
|
|
HO |
O |
CH2 |
CH |
COOH |
|
|
|
NH2 |
|
I
Трийодотиронін (рТ3) неактивний
Монодейодування зовнішнього кільця молекули тироксину приводить до активації гормону, дейодування внутрішнього кільця — до утворення неактивної форми — рТ3.
Протеїнзв’язаний тироксин є сховищем гормону в крові. Тільки вільний тироксин і Т3 можуть без перешкод переноситися через мембрану в цитоплазму клітини-мішені, де тироксин ферментативно перетворюється на активніший Т3. У
клітинному ядрі Т3 зв’язується зі специфічними рецепторами, які належать до сімейства ядерних рецепторів, з’єднаних із хроматином. Ці рецептори неактивні, доки Т3 не дифундує з цитоплазми. Взаємодія Т3 з цим рецептором підвищує швидкість транскрипції РНК, отже, збільшується продукція різних білків (рис. 15.3). Тиреоїдні гормони мають також посттранскрипційні ефекти. Тиреоїдні гормони найсильніше впливають на поділ і диференціювання клітин та енергетичний обмін організму. Зміни в енергетичному обміні зовні виражаються в підвищеному споживанні кисню і продукуванні теплоти. Йодотироніни діють на обмін речовин
удвох різних напрямках:
—через ядерні рецептори на хромосоми ядра. Гормони є індукторами синтезу більше 100 різних ферментів. У клітині під дією йодотиронінів зростає кількість мітохондрій, а в багатьох із них збільшуються в розмірах кристи, які містять велику кількість дихальних ланцюгів;
—через активацію йодотиронінами аденілатциклази в тканинах і збільшення вмісту цАМФ, яка активує ліполіз у жировій тканині й глікогеноліз у печінці та м’язах.
Внутрішньоклітинні механізми метаболічних ефектів щитоподібної залози потребують підвищеної активності Na+/К+-АТФази. Посилення активності цього ферменту можливе тільки при високому рівні АТФ, утилізація якої супроводжується утворенням тепла. Підвищена потреба в АТФ забезпечується процесом окисного фосфорилування в мітохондріях. Йодотироніни роз’єднують дихання і фосфорилування, через що зростає вільне окиснення і збільшується теплопродукція.
Тироксинзв’я- |
|
|
Цитоплазма |
ДНК |
|
|
|
зувальний |
Ядро |
|
|
|
|
||
глобулін |
|
|
|
|
|
Т3 |
мРНК |
|
|
|
|
|
|
|
мРНК |
Т4 |
|
Т3 |
Т3 |
|
|
|
|
|
|
|
Синтез |
|
|
|
білка |
Т4 |
Т4 |
Т3 |
|
|
Зв’язувальний |
|
|
|
|
білок |
|
Кров |
|
Клітина-мішень |
|
Рис. 15.3. Механізм дії тиреоїдного гормону |
|||
|
на клітину-мішень |
|
|
207
|
|
|
|
Порушення функції щитоподібної залози |
дження чи в ранньому дитинстві, відомий як кре- |
|||||||||||||||
|
|
|
|
|
|
Гіпертиреоз |
|
|
|
тинізм. При гіпофункції щитоподібної залози в |
||||||||||
|
|
|
|
|
|
|
|
|
дорослих розвивається мікседема. |
|||||||||||
|
|
|
Гіпертиреоз може виникнути при: |
|||||||||||||||||
|
|
|
Лікування гіпотиреозу полягає в призначенні |
|||||||||||||||||
|
|
|
— надмірній стимуляції залози, як при хворобі |
пероральної замісної терапії тиреоїдним гормо- |
||||||||||||||||
|
Грейвса (базедова хвороба, тиреотоксикоз); |
ном, при первинному гіпотиреозі (найчастіша |
||||||||||||||||||
|
|
|
— автономній секреції тироксину, як при ток- |
причина) — нормалізації вмісту тирозинстиму- |
||||||||||||||||
|
сичній аденомі та токсичному багатовузловому |
лювального гормону. |
||||||||||||||||||
|
зобі; |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
— надлишковому прийомі екзогенного гормо- |
|
||||||||||||||||
|
ну. |
|
|
|
|
|
|
|
|
|
|
|
|
|
Гормональне регулювання метаболізму |
|||||
|
|
|
При надмірному вмісті тиреоїдного гормону |
кальцію та фосфору |
||||||||||||||||
|
зростає потреба клітин у кисні, збільшується про- |
|||||||||||||||||||
|
|
|||||||||||||||||||
|
дукція тепла і використання метаболітів, що зви- |
Постійний рівень Са в крові дуже важливий |
||||||||||||||||||
|
чайно призводить до зниження маси тіла. Озна- |
для м’язової діяльності, збудливості нервової тка- |
||||||||||||||||||
|
ками тиреотоксикозу є схуднення, підвищення |
нини, для скоротливої здатності міокарда, при |
||||||||||||||||||
|
температури тіла, інтоксикація недоокисненими |
згортанні крові. |
||||||||||||||||||
|
продуктами обміну, ураження серцевої та нерво- |
У регулюванні метаболізму кальцію беруть |
||||||||||||||||||
|
вої систем. |
|
|
|
|
|
|
|
|
|
|
|
|
|
участь три гормони: паратгормон, кальцитонін, |
|||||
|
|
|
Надлишкова секреція тирозинстимулюваль- |
1,25-дигідроксихолекальциферол. Четвертий гор- |
||||||||||||||||
|
ного гормону (ТСГ) дуже рідко стає причиною |
мон — пептид, подібний до паратгормону (PTHrP) |
||||||||||||||||||
|
тиреотоксикозу. |
|
|
|
|
|
|
|
|
|
|
|
|
|
(від англ. pаrаthyrеoid hormones — related pro- |
|||||
|
|
|
Лікування гіпертиреозу (гіпертироксинемії) |
stein), він діє на один із рецепторів паратгормо- |
||||||||||||||||
|
залежить від його причини і полягає у призна- |
ну і є важливим для розвитку скелетних м’язів in |
||||||||||||||||||
|
ченні антитиреоїдних препаратів (метимазолу, |
utero. На метаболізм кальцію також можуть |
||||||||||||||||||
|
пропілтіоурацилу), β -блокаторів (пригнічують |
впливати гормони, що регулюють вміст фосфору, |
||||||||||||||||||
|
перетворення Т4 на активну форму Т3), радіоак- |
глюкокортикоїди, гормон росту, естрогени та |
||||||||||||||||||
|
тивного йоду, проведенні хірургічної операції |
різноманітні фактори росту. |
||||||||||||||||||
|
(рис. 15.4). |
|
|
|
|
|
|
|
|
|
|
|
|
|
Кальцій у кістках буває двох типів: легкооб- |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
мінний пул і набагато більший пул стабільного |
I |
|
|
|
|
|
|
|
|
T4 |
|
|
T3 |
|
|
|
Метаболічні |
кальцію, здатний лише до повільного обміну. Є |
|||
|
|
|
|
|
|
|
|
|
|
|
ефекти |
дві різні незалежні гомеостатичні системи, що |
||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
взаємодіють і впливають на кальцій у кістках. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Одна система регулює Са2+ у плазмі, за її до- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Симптоми |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
помогою щодня 500 ммоль Са2+ потрапляє в |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
кістковий пул кальцію, здатний до легкого об- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
міну. |
|
|
Пропілтіоурацил, |
|
|
|
β -блокатори блокують ак- |
|
|||||||||||||
|
метимазол блокують |
|
|
|
тивацію тиреоїдного гормо- |
|
Друга система пов’язана з ремоделюванням |
|||||||||||||
|
|
вбудовування йоду |
|
|
|
ну і зменшують симптоми |
|
кістки шляхом постійної взаємодії між резорбцією |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
та утворенням кістки, у дорослих вона на 95 % |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
відповідає за утворення кістки. Проте взаємо- |
|
|
|
|
|
Йодид літію блокує |
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
обмін Са2+ між плазмою та цим стабільним пулом |
||||||||||
|
|
|
|
|
вивільнення гормонів |
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
становить 7,5 ммоль за добу. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 15.4. Лікування гіпертиреозу |
Значну кількість кальцію фільтрують нирки, |
|||||||||||||||
|
|
|
|
однак 98–99 % його реабсорбується (близько |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
60 % у проксимальних канальцях, решта — у |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
висхідній частині петлі Генле та дистальних ка- |
|
|
|
|
|
|
Гіпотиреоз |
|
|
|
нальцях нефрону). Реабсорбцію в дистальних |
||||||||||
|
|
|
Гіпотиреоз може виникнути при: |
|
|
|
канальцях регулює паратгормон. |
|||||||||||||
|
|
|
|
|
|
Паратгормон (паратиреоїдний гормон, полі- |
||||||||||||||
|
|
|
— недостатності щитоподібної залози (пер- |
|||||||||||||||||
|
|
|
пептид) — його секретують паращитоподібні за- |
|||||||||||||||||
|
винний гіпотиреоз, |
|
чи автоімунний тиреоїдит |
|||||||||||||||||
|
Хашимото); |
|
|
|
|
|
|
|
|
|
|
|
|
|
лози. Існує три типи рецепторів гормону. Один |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
із них зв’язує як паратгормон, так і пептид, по- |
||||||
|
|
|
— недостатності гіпофіза (вторинний гіпоти- |
|||||||||||||||||
|
реоз) чи гіпоталамуса (третинний гіпотиреоз); |
дібний до паратгормону, і через GS активує аде- |
||||||||||||||||||
|
|
|
— деяких рідких станах, таких як недостат- |
нілатциклазу. Цей рецептор також може за до- |
||||||||||||||||
|
ність йоду і спричинена лікарськими препарата- |
помогою білка G активувати фосфатидилінози- |
||||||||||||||||||
|
q |
|||||||||||||||||||
|
толову систему. Білок з активністю паратгормо- |
|||||||||||||||||||
|
ми недостатність щитоподібної залози. |
ну утворюється в багатьох тканинах. У хрящах, |
||||||||||||||||||
|
|
|
При дефіциті гормонів щитоподібної залози |
|||||||||||||||||
|
утворення тепла і використання кисню зменшу- |
які розвиваються, він стимулює проліферацію |
||||||||||||||||||
|
ються, а метаболіти окиснюються повільніше, у |
хондроцитів і пригнічує їх мінералізацію. |
||||||||||||||||||
|
результаті чого маса тіла збільшується (гіпоти- |
Механізм дії паратгормону |
||||||||||||||||||
|
реоз). Виникають набряки, зумовлені збільшен- |
|||||||||||||||||||
|
1. Його головна функція — мобілізація каль- |
|||||||||||||||||||
|
ням гідрофільності тканин (слизовий набряк). |
|||||||||||||||||||
|
Гіпотиреоз, який виявляється від самого наро- |
цію з кісток і збільшення екскреції фосфату з се- |
||||||||||||||||||
208