Біологічна та біоорганічна хімія_Мардашко О.О._ изд. 2008-342 с._ОНМедУ-2012
.pdf

 HO
HO 

 O
O

 O
O
 O
O
|
CH2OH |
|
|
O CH2OH |
Жіночі статеві гормони (естрогени) утворю- |
|||||||
|
C O |
|
|
HC C O |
ються переважно у фолікулах яєчників і в жов- |
|||||||
|
HO |
ОН |
|
HO |
|
тому тілі. У фолікулах яєчників синтезуються три |
||||||
|
|
|
|
|
|
гормони: естрон (фолікулін), естрадіол і естріол. |
||||||
|
|
|
|
|
|
У жовтому тілі синтезується прогестерон. |
|
|||||
O |
Кортизол |
|
O |
Альдостерон |
Основним естрогеном, який секретується, є |
|||||||
|
естрадіол, інші естрогени (естрон і естріол) — про- |
|||||||||||
|
|
|
|
|
|
дукти його перетворення. |
|
|
|
|||
|
|
|
|
|
|
H C |
O |
|
|
OHOH |
||
|
|
|
|
|
|
|
|
H C |
|
|||
|
|
|
|
|
|
3 |
|
|
H3C3 |
|
CH3 OH |
|
11-β -гідроксистероїддегід- |
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
||||
рогеназа в ендоплазма- |
|
|
|
|
|
|
|
|
|
|||
|
тичному ретикулумі |
Мембрана |
|
HO |
|
|
HO |
|
|
|||
|
|
CH2OH |
|
клітини |
|
|
|
|
|
|||
|
|
|
|
Естрон (у фолікулах) |
Естрадіол (у фолікулах) |
|||||||
|
|
C O |
|
|
|
|||||||
|
O |
OH |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H C OH |
|
|
|
CH3 |
||
|
|
|
|
|
|
|
H C C O |
|||||
|
|
|
|
|
|
3 |
|
OH |
||||
|
O |
|
|
|
|
|
|
3 |
|
|
||
|
|
|
|
|
|
|
|
|
|
|||
|
Кортизон |
|
|
|
|
|
|
|
H3C |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
HO |
|
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
Естріол |
|
|
Прогестерон |
|||
|
|
|
|
|
|
(у фолікулах) |
|
|
(у жовтому тілі) |
|||
|
|
МР |
|
|
|
Всі естрогени, що синтезуються у фолікулах, |
||||||
|
|
|
|
|
мають високий ступінь ненасиченості кільця А |
|||||||
|
|
|
|
|
|
|||||||
|
|
|
Ядро |
|
(ароматичність) і 18 атомів Карбону. Продукція |
|||||||
|
|
|
|
естрогенів має циклічний характер. У першу |
||||||||
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
фазу менструального циклу — овуляцію — фолі- |
||||||
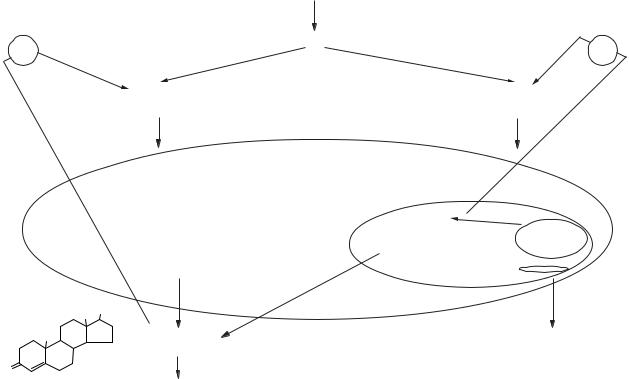
Рис. 15.18. Механізм дії альдостерону на клітину |
кулами продукується, головним чином, естраді- |
|||||||||||
ол, що забезпечує визрівання яйцеклітини. У дру- |
||||||||||||
|
(МР — мінералокортикоїдний рецептор) |
|||||||||||
|
гу фазу — функціонування жовтого тіла — про- |
|||||||||||
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
дукується прогестерон, який стимулює розвиток |
||||||
Гормони статевих залоз |
|
|
матки, розвиток заплідненого яйця і плода. |
|||||||||
|
|
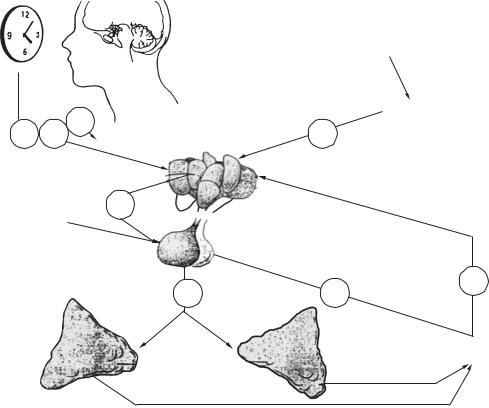
На функцію статевих залоз і продукцію ста- |
||||||||||
|
|
|
|
|
|
|||||||
Статеві залози продукують статеві гормони і |
тевих гормонів впливає гіпоталамо-гіпофізарна |
|||||||||||
статеві клітини. Деяка кількість статевих гор- |
система. Фоліберин, люліберин гіпоталамуса ак- |
|||||||||||
монів утворюється, крім того, у плаценті (під час |
тивують продукцію фолітропіну і лютропіну |
|||||||||||
вагітності) і корі надниркових залоз. Статеві гор- |
гіпофіза, які, у свою чергу, через систему цикліч- |
|||||||||||
мони синтезуються із загального попередника — |
них нуклеотидів стимулюють синтез гормонів і |
|||||||||||
холестеролу, багато стадій їхнього утворення |
клітин статевих залоз. У жінок фолітропін стиму- |
|||||||||||
збігаються, тому незначна кількість жіночих і чо- |
лює дозрівання фолікулів і синтез естрогенів, а |
|||||||||||
ловічих статевих гормонів синтезується в осіб |
лютропін — функціонування жовтого тіла і син- |
|||||||||||
протилежної статі (див. рис. 15.15). Оскільки ста- |
тез прогестерону. У чоловіків фолітропін стиму- |
|||||||||||
теві гормони зумовлюють формування вторин- |
лює проліферацію сперматогенного епітелію, а |
|||||||||||
них ознак відповідної статі, то від кількості гор- |
лютропін — синтез тестостерону. Статеві залози, |
|||||||||||
монів протилежної статі залежить виразність про- |
як і кора надниркових залоз, перебувають із гіпо- |
|||||||||||
явів статевої приналежності людини. Розвиток |
таламо-гіпофізарною системою у реципрокних |
|||||||||||
статі визначається статевими хромосомами й |
відносинах (від англ. reciprocal — взаємний). |
|||||||||||
особливостями секреції статевих гормонів гона- |
Біологічна дія статевих гормонів |
|
||||||||||
дами в ембріональний період. Чоловічі статеві |
|
|||||||||||
|
|
|
|
|
|
|||||||
гормони — андрогени (від грецьк. andros — чо- |
Як і для гормонів кори надниркових залоз, |
|||||||||||
ловік). Характерною ознакою андрогенів є те, |
для статевих гормонів у цитоплазмі клітин-міше- |
|||||||||||
що вони мають 19 атомів Карбону. До чоловічих |
ней є спеціальні білки-рецептори, що взаємодіють |
|||||||||||
статевих гормонів належать тестостерон і 5-α -ди- |
із молекулою гормону з утворенням гормон-ре- |
|||||||||||
гідротестостерон: |
|
|
|
|
цепторного комплексу. У цьому комплексі гормон |
|||||||
|
H C OH |
|
|
OH |
транспортується до ядерної мембрани, проникає |
|||||||
|
|
H3C |
в ядро й зв’язується зі специфічними акцепторни- |
|||||||||
|
3 |
|
|
CH3 |
||||||||
|
|
|
|
|
ми ділянками хроматину, впливаючи на гене- |
|||||||
|
H3C |
|
|
|
|
|||||||
|
|
|
|
|
тичний апарат клітини. |
|
|
|
||||
|
|
|
|
|
|
|
|
|
||||
|
O |
|
O |
|
|
Андрогени й естрогени підсилюють біосинтез |
||||||
|
|
|
|
інформаційної, транспортної й рибосомальної |
||||||||
|
|
|
5-α -Дигідротестостерон |
|||||||||
|
Тестостерон |
|
РНК, активуючи амінотрансферази (амінування |
|||||||||
|
|
|
|
|
|
223 |
|
|
|
|
|
|