

Тема 7 динамика численности популяций
Объектом изучения демоэкологии, или популяционной экологии, служит популяция. Ее определяют как группу организмов одного вида (внутри которой особи могут обмениваться генетической информацией), занимающую конкретное пространство и функционирующую как часть биотического сообщества. Популяция характеризуется рядом признаков; единственным их носителем является группа, но не особи в этой группе. Важнейшее свойство популяции — плотность, т. е. число особей, отнесенное к некоторой единице пространства.
7.1. Экспоненциальная и логистическая кривые роста
Численность популяции определяется в основном двумя противоположными явлениями — рождаемостью и смертностью. Пусть N — численность популяции. Тогда отношение dN/dt означает мгновенную скорость изменения N, т. е. изменение N в момент времени t, а отношение dN/dt N удельную мгновенную скорость изменения численности популяции. Так, в популяции инфузорий с исходной численностью 100 особей и с численностью, равной 200 особей через час, рост популяции равен 100 особей в час, а рост популяции в расчете на одну особь равен 100/100-1 = 1 особь в час.
Таким же способом определяют рождаемость и смертность. Если dN - число рожденных особей в популяции, то коэффициент рождаемости b = dN/dt, если dN — число погибших особей, то коэффициент смертности d = dN/dt. Разность r = b-d представляет собой коэффициент прироста изолированной популяции, в которой нет ни эмиграции, ни иммиграции. Коэффициент прироста популяции—не что иное, как врожденная (специфическая) скорость естественного увеличения популяции, называемая иногда мальтузианским параметром. Эта величина является мерой мгновенной удельной (в расчете на одну особь)
116
скорости изменения размера популяции и выражается как число особей, отнесенное к единице времени в расчете на одну особь. Различают максимальную мгновенную скорость прироста популяции (rmax) и фактическую скорость увеличения популяции ra=b-d. При воображаемых идеальных условиях, когда значение b максимально, a d минимально, га достигает наибольшей величины — rmax
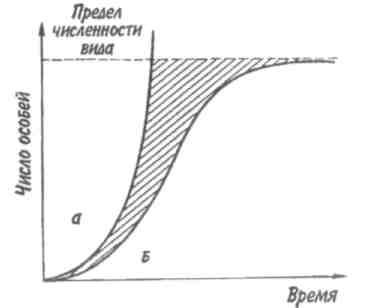
Рис. 7.1. Экспоненциальная (а) и логистическая (б)
кривые роста популяции.
Заштрихованная площадь — сопротивление среды.
Если допустить, что коэффициент прироста имеет постоянную величину, то зависимость численности популяции от времени будет выражаться экспоненциальной кривой (рис. 7.1,а). Если r = const, то dN/dt = rN, следовательно, если N0 — численность популяции в исходный момент, Nt— ее численность в момент t, то Nt = N0ert. Экспоненциальная кривая выражает так называемый биотический потенциал.
В природе в основном наблюдается иная картина. Прежде всего коэффициент прироста не остается постоянным, так как рождаемость и смертность меняются в зависимости от условий среды и возраста организмов, а пища и территория редко предоставлены в достаточном объеме. Чаще всего реальный рост численности популяции выражается S-образной зависимостью, которую называют логистической кривой роста (рис. 7.1,6). Урав-
117
нение логистической кривой отличается от уравнения биотического потенциала корректирующим фактором: (K-N)/ К, где К— максимальное число особей, способных жить в рассматриваемой среде, т. е. асимптота кривой. Отсюда математическое выражение для логистической кривой имеет вид: dN/dt = rN[(K-N)K-1] или Nt = K/1+ea-rt, где a = r/ К. Пространство, заключенное между биотическим потенциалом и логистической кривой роста, представляет собой сопротивление среды.