 волокна идут прямо к коре, минуя
волокна идут прямо к коре, минуя  Проекционные волокна между корой и внутренней капсулой образуют
Проекционные волокна между корой и внутренней капсулой образуют
лучистый, или сияющий венец (corona radiata). Зрительная лучистость, или сияние, соединяющее боковое коленча5 тое тело с 17 полем, и слуховое сияние, соединяющее срединное коленчатое те5 ло с 41 полем, также являются частью проекционной системы (см. рис. 1.20, 2.2, 3.12, 3.37, 8.14 и 
Ассоциативные волокна
Эти волокна составляют основу белого вещества. Они соединяют как близле5 жащие, так и отдаленные корковые об5 ласти. Таким способом все функцио5 нально значимые корковые области мо5 гут тесно взаимодействовать, позволяя тем самым мозговой коре выполнять ее ассоциативные и интегративные функции. Обширные связи между от5 дельными корковыми областями слу5 жат вероятным объяснением того факта, что потеря мозговой функции вследст5 вие очагового повреждения не всегда постоянна. Возможно, некоторые волок5 на остаются интактными и могут вос5 становить часть утраченных функций после упражнения в течение определен5 ного времени.
Наиболее важные пучки ассоциатив5 ных волокон показаны на рис. 8.14 и 8.15. Некоторые из них заслуживают отдельного разговора.
Аркообразные волокна называют также U5образными волокнами. Они за5 нимают верхние подкорковые отделы белого вещества, формируя U5образный слой вокруг глубинных впадин коры и соединяя соседние корковые поля. Из5 вестна их резистентность при некото5 рых демиелинизирующих лейкоэнце5 фалопатиях.
Верхний продольный пучок проходит кзади от островка в передне5заднем на5 правлении и соединяет лобные доли с теменно5затылочными и височными
извилинами. Его лобновисочная порция огибает задний край сильвиевой 
и называется аркообразным пучком (fas$  ciculus arcuatus). Предполагают, что этот пучок соединяет височные (Вернике) и лобные (Брока) речевые центры. Ниж$ ний продольный пучок соединяет височ5 ные и затылочные доли.
ciculus arcuatus). Предполагают, что этот пучок соединяет височные (Вернике) и лобные (Брока) речевые центры. Ниж$ ний продольный пучок соединяет височ5 ные и затылочные доли.  пучок огибает передние отделы сильвие5 вой щели, соединяя орбитальную долю
пучок огибает передние отделы сильвие5 вой щели, соединяя орбитальную долю
с полюсом височной. Пояс (cingulum) является одним из ассоциативных пуч5 ков лимбической системы и, огибая мо5 золистое тело на всем его протяжении, соединяет подмозолистую область с па5 рагиппокамповой извилиной. Очень важными ассоциативными волокнами являются верхние и нижние  лобные пучки и вертикальный затылоч$ ный пучок.
лобные пучки и вертикальный затылоч$ ный пучок.
волокна
Мозолистое тело (corpus callosum) яв5 ляется самой большой  но5 вой коры. Волокна проходят через эту комиссуру из одного полушария в дру5 гое и затем расходятся в разных на5 правлениях, соединяя все гомотопиче5 ские области коры за исключением пер5 теичной зрительной коры (поле 17) и проекций руки и ноги в соматосенсор5 ной
но5 вой коры. Волокна проходят через эту комиссуру из одного полушария в дру5 гое и затем расходятся в разных на5 правлениях, соединяя все гомотопиче5 ские области коры за исключением пер5 теичной зрительной коры (поле 17) и проекций руки и ноги в соматосенсор5 ной  мозолистой лучис5 тости пересекают волокна лучистого венца (corona radiata) и других ассоциа5 тивных пучков. Поскольку мозолистое тело короче больших полушарий, во5 локна идущие через клюв, колено или валик мозолистого тела дугообразно из5 гибаются в направлении лобных и, со5 ответственно, затылочных полюсов. Эти дугообразные пучки волокон называют5 ся передними (малыми) (forceps fronta$
мозолистой лучис5 тости пересекают волокна лучистого венца (corona radiata) и других ассоциа5 тивных пучков. Поскольку мозолистое тело короче больших полушарий, во5 локна идущие через клюв, колено или валик мозолистого тела дугообразно из5 гибаются в направлении лобных и, со5 ответственно, затылочных полюсов. Эти дугообразные пучки волокон называют5 ся передними (малыми) (forceps fronta$  изадними(большими)(forcepsocci$
изадними(большими)(forcepsocci$  щипцами мозолистой лучистос5 ти (см. рис. 8.15в). Передняя комиссура
щипцами мозолистой лучистос5 ти (см. рис. 8.15в). Передняя комиссура  anterior) является спайкой между обонятельным мозгом, височны5 ми долями и, в меньшей степени, над5
anterior) является спайкой между обонятельным мозгом, височны5 ми долями и, в меньшей степени, над5


 с поверхности коры.
с поверхности коры. Первый или молекулярный
Первый или молекулярный  или наружный гранулярный слой:
или наружный гранулярный слой: 
 волок5 на. Отходящие от верхнего края клеток дендриты простираются в первый мо5 лекулярный слой. Остальные дендриты ветвятся в основном в наружном пира5 мидном слое.
волок5 на. Отходящие от верхнего края клеток дендриты простираются в первый мо5 лекулярный слой. Остальные дендриты ветвятся в основном в наружном пира5 мидном слое. или внутренний грану лярный слой:
или внутренний грану лярный слой: 


 ный слой:
ный слой: 
 8.12 Упрощенная схема не* которых
8.12 Упрощенная схема не* которых 

 цепей
цепей  Lorento de No и Larsell).
Lorento de No и Larsell). черным цветом. Они исходят из таламуса в виде проекционных во5 локон (указаны на рисунке цифрой) и из других участков коры виде проекци5 онных волокон (цифра 2) и заканчива5 ются на гранулярных клетках второго и четвертого слоя коры. Эфферентные волокна показаны красным цветом. Од5 ни аксоны пирамидных клеток пятого слоя идут через внутреннюю капсулу к таламусу, полосатому телу, ядрам ство5 ла и спинному мозгу
черным цветом. Они исходят из таламуса в виде проекционных во5 локон (указаны на рисунке цифрой) и из других участков коры виде проекци5 онных волокон (цифра 2) и заканчива5 ются на гранулярных клетках второго и четвертого слоя коры. Эфферентные волокна показаны красным цветом. Од5 ни аксоны пирамидных клеток пятого слоя идут через внутреннюю капсулу к таламусу, полосатому телу, ядрам ство5 ла и спинному мозгу  другие явля5 ются ассоциативными и комиссураль5 ными волокнами и связаны с прочими корковыми областями
другие явля5 ются ассоциативными и комиссураль5 ными волокнами и связаны с прочими корковыми областями 
 кора является
кора является  Су5 ществуют корковые поля мозга взрос5 лого, где эти шесть слоев трудно раз5
Су5 ществуют корковые поля мозга взрос5 лого, где эти шесть слоев трудно раз5
 органов чувств (зрения, слуха), гра5 нулярные клетки преобладают над пи5 рамидными. Поэтому, говорят о
органов чувств (зрения, слуха), гра5 нулярные клетки преобладают над пи5 рамидными. Поэтому, говорят о  поля 4 и 6. Бродман выделил немногим более 50 различ5 ных архитектонических полей. Обычно пользуются составленной им картой, потому что она проще карты фон Эко5 номо, которая насчитывает вдвое боль5 ше полей. Некоторые исследователи вы5 деляют более двухсот различных полей. Von Economo (1925) обнаружил, что различные архитектонические поля имеют ту или иную общую черту и могут быть сведены к пяти основным типам, изображенным на рис. 8.13. Как видно на рис. 8.9, цитоархитектоничес5 кие поля точно не соответствуют ходу извилин. Они частично перекрываются и подвержены индивидуальным вариа5
поля 4 и 6. Бродман выделил немногим более 50 различ5 ных архитектонических полей. Обычно пользуются составленной им картой, потому что она проще карты фон Эко5 номо, которая насчитывает вдвое боль5 ше полей. Некоторые исследователи вы5 деляют более двухсот различных полей. Von Economo (1925) обнаружил, что различные архитектонические поля имеют ту или иную общую черту и могут быть сведены к пяти основным типам, изображенным на рис. 8.13. Как видно на рис. 8.9, цитоархитектоничес5 кие поля точно не соответствуют ходу извилин. Они частично перекрываются и подвержены индивидуальным вариа5
 агранулярная; 2
агранулярная; 2  лобная; 3
лобная; 3  теменная; 4
теменная; 4  полюсная; 5
полюсная; 5  гранулярная (кониокортес). а распределение типов мозговой коры по конвексу; б их цитоархитектурный
гранулярная (кониокортес). а распределение типов мозговой коры по конвексу; б их цитоархитектурный
 организации мозговой ко5 ры, рассмотрим ее связи.
организации мозговой ко5 ры, рассмотрим ее связи. двусторонних проводов,
двусторонних проводов,  кору и подкорковые цент5
кору и подкорковые цент5 (проекционные волокна),
(проекционные волокна),  superior
superior пирамидной системах. Они являются волокнами корковоядерного, корково5
пирамидной системах. Они являются волокнами корковоядерного, корково5 136
136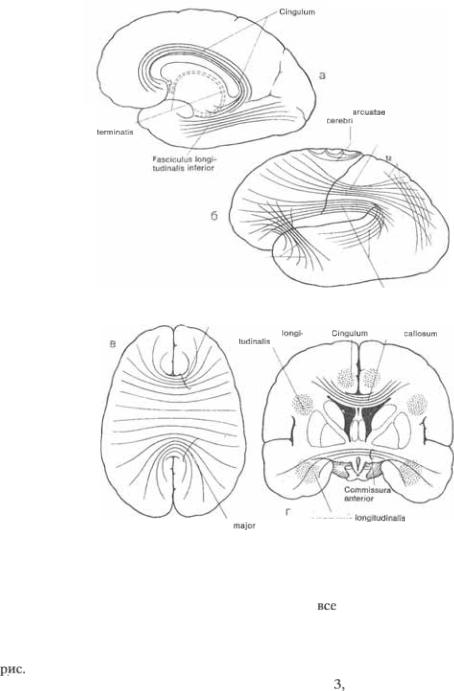
 8.15Схематическое изображе* ние основных ассоциативных и
8.15Схематическое изображе* ние основных ассоциативных и 
 пучков волокон.
пучков волокон.
 superior
superior
 verticalis
verticalis et Fasciculus arcuatus
et Fasciculus arcuatus
 лобной корой.
лобной корой. 
 или
или

 седалищем ду5 ши и
седалищем ду5 ши и  функции коры должны были быть нераздельны. Эта теория считалась общепринятой после того как Пьер Флорен, сравнительный анатом и физиолог из Парижского университета опубликовал ее в 1824 г., исходя из своих опытов на
функции коры должны были быть нераздельны. Эта теория считалась общепринятой после того как Пьер Флорен, сравнительный анатом и физиолог из Парижского университета опубликовал ее в 1824 г., исходя из своих опытов на  (в
(в  на птицах).
на птицах). также убедитель5 но показал доминантность одного из полушарий. В США Вейр Митчелл из Филадельфии (1860) отметил, что мышцы на одной стороне тела иннер5 вируются корой противоположного по5 лушария головного мозга. Его опыты
также убедитель5 но показал доминантность одного из полушарий. В США Вейр Митчелл из Филадельфии (1860) отметил, что мышцы на одной стороне тела иннер5 вируются корой противоположного по5 лушария головного мозга. Его опыты предста5 вительство в коре контрлатеральных движений. Эти опыты были продолже5 ны другими выдающимися физиолога5 ми, такими как Ферриер (1876), Бивор
предста5 вительство в коре контрлатеральных движений. Эти опыты были продолже5 ны другими выдающимися физиолога5 ми, такими как Ферриер (1876), Бивор (1906). Что касается сен5 сорного коркового представительства, фон Гудден (1870) показал, что удале5 ние обох глаз у молодого животного приводило к задержке развития заты5 лочных долей. Мунк (1979) установил, что собаки без затылочных долей яв5 ляются слепыми и что повреждение только коры поверхности затылочной доли вызывает
(1906). Что касается сен5 сорного коркового представительства, фон Гудден (1870) показал, что удале5 ние обох глаз у молодого животного приводило к задержке развития заты5 лочных долей. Мунк (1979) установил, что собаки без затылочных долей яв5 ляются слепыми и что повреждение только коры поверхности затылочной доли вызывает  назы5 ваемую теперь
назы5 ваемую теперь 

 8.16 Цитоархитектоническая локализация функций в коре большого мозга (по К. Клейсту).
8.16 Цитоархитектоническая локализация функций в коре большого мозга (по К. Клейсту).  левого полушария.
левого полушария.
 тела)
тела) 8.17 Цитоархитектоническая локализация функций в коре большого мозга (по К Клейсту). Медиальная поверхноаь правого полушария. (Рис. 8.16 и 8.17 взяты из: К. Kleist Gehirnpathologie.
8.17 Цитоархитектоническая локализация функций в коре большого мозга (по К Клейсту). Медиальная поверхноаь правого полушария. (Рис. 8.16 и 8.17 взяты из: К. Kleist Gehirnpathologie.  Handbuch der
Handbuch der  ErfahrungenimWeltkrieg1914/18,Bd.IV,Barth,Leipzig,1922*1934).
ErfahrungenimWeltkrieg1914/18,Bd.IV,Barth,Leipzig,1922*1934). 1909;
1909;  Vogt, 1910; V. Economo и
Vogt, 1910; V. Economo и  1925; Beck, 1925; Rose, 1928).
1925; Beck, 1925; Rose, 1928).  также заинтере5 совался тем обстоятельством, что обра5 зующие кору клеточные слои имеет раз5 личное функциональное значение
также заинтере5 совался тем обстоятельством, что обра5 зующие кору клеточные слои имеет раз5 личное функциональное значение  1908;
1908;  1911; Kappers, 1920 и мно5 гие другие).
1911; Kappers, 1920 и мно5 гие другие).
 синергия контрлатеральной
синергия контрлатеральной ноги.
ноги. 0.
0.  0. Foerster (Eds), Handbuch der Neurologie, Bd. IV, Springer, Berlin,
0. Foerster (Eds), Handbuch der Neurologie, Bd. IV, Springer, Berlin,