

Дуус / Топический диагноз в неврологии
.PDF178 3 Ствол мозга
женной гипертонии мышц5разгибате5 лей головы и конечностей, сохраняю5 щейся до тех пор, пока остаются ин5 тактными ростральные и латеральные части ретикулярной формации и ядра вестибулярных нервов.
Наиболее частыми односторонними  объемными па5 тологическими процессами являются внутримозговые гематомы естественого происхождения (разрыв пораженных в результате заболевания артерий или со5 судистых мальформаций); нарастаю5 щий на протяжение первых одного— шести дней очаг обширного инфаркта; опухоли, а также травматические экс5 традуральные, субдуральные и внутри5 мозговые гематомы. Двустороннее сдав5 ление среднего мозга наблюдается обычно при увеличении в объеме обоих полушарий головного мозга. Наиболее частой причиной генерализованного на5 бухания мозга является инсульт, вне зависимости от его природы.
объемными па5 тологическими процессами являются внутримозговые гематомы естественого происхождения (разрыв пораженных в результате заболевания артерий или со5 судистых мальформаций); нарастаю5 щий на протяжение первых одного— шести дней очаг обширного инфаркта; опухоли, а также травматические экс5 традуральные, субдуральные и внутри5 мозговые гематомы. Двустороннее сдав5 ление среднего мозга наблюдается обычно при увеличении в объеме обоих полушарий головного мозга. Наиболее частой причиной генерализованного на5 бухания мозга является инсульт, вне зависимости от его природы.
При объемных патологических про5 цессах в задней черепной ямке, таких, как гематома мозжечка (см. рис. 4.10), может произойти смещение вершины горки червя мозжечка и окружающих ее тканей в супратенториальное про5 странство (см. рис. 4.11). Она выталки5 вает средний мозг в вентральном на5 правлении, к базальной цистерне. Такое заднее вклинение не угрожает глазо5 двигательным нервам; следовательно, отсутствуют зрачковые расстройства, описанные выше как результат сдавле5
 3.72 Децеребрационная
3.72 Децеребрационная 
 с
с  спаз* мами.
спаз* мами.
ния этих нервов. Однако вместо них могут наблюдаться другие нарушения движений глазных яблок, особенно если имеется также и давление на покрышку варолиева моста. Не характерны ранняя утрата сознания и децеребрационная ригидность. Могут быть пирамидные знаки, связанные с ущемлением мин5 далин мозжечка в большом затылочном отверстии. Когда миндалины мозжечка смещаются в заднюю часть большого затылочного отверстия, они часто при5 жимают продолговатый мозг к передне5 му краю большого затылочного отверс5 тия, подвергая локальной компрессии пирамиды. Ипсилатеральная по отно5 шению к очагу в мозжечке пирамида может сдавливаться сильнее, чем дру5 гая, что обуславливает асимметрию двигательных симптомов. Кроме того, возможна гипестезия или анестезия на одной или обеих половинах туловища. При остро возникающих мозжечковых поражениях, таких, как гематома, име5 ется значительная угроза развития ды5 хательного паралича. В отличие от ды5 хательных нарушений при компрессии среднего мозга, остановка дыхания при поражении продолговатого мозга часто внезапна и неожиданна.
Очевидно, что при наличии вкли5 нения в вырезку намета мозжечка и большое затылочное отверстие, выпол5 нение люмбальной пункции чрезвычай5 но опасно и может спровоцировать рез5 кое, фатальное ухудшение состояния больного.

 синдром
синдром
Термин «апаллический» 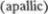 обозна5 чает утрату функции pallium, то есть плаща головного мозга — мозговой ко5 ры. Возможно, что за развитие первой фазы так называемого апаллического синдрома ответственно повреждение среднего мозга, возникающее при тен5 ториальном вклинении. Продолжитель5 ная глубокая кома после тяжелого по5 вреждения головы предполагает суще5 ствование компрессии среднего мозга, которое порождает дополнительные симптомы в виде спазмов сгибателей или разгибателей, ригидности, глазо5 двигательных нарушений, односторон5 них либо двусторонних пирамидных симптомов. После окончания этой фазы может пройти несколько недель до по5 явления симптомов генерализованного поражения головного мозга. Глубокая кома переходит в «бодрствующую» кому,
обозна5 чает утрату функции pallium, то есть плаща головного мозга — мозговой ко5 ры. Возможно, что за развитие первой фазы так называемого апаллического синдрома ответственно повреждение среднего мозга, возникающее при тен5 ториальном вклинении. Продолжитель5 ная глубокая кома после тяжелого по5 вреждения головы предполагает суще5 ствование компрессии среднего мозга, которое порождает дополнительные симптомы в виде спазмов сгибателей или разгибателей, ригидности, глазо5 двигательных нарушений, односторон5 них либо двусторонних пирамидных симптомов. После окончания этой фазы может пройти несколько недель до по5 явления симптомов генерализованного поражения головного мозга. Глубокая кома переходит в «бодрствующую» кому,
характеризующуюся непродолжитель5 ными периодами бодрствования. Боль5 ной открывает глаза, но не реагирует на окружающие стимулы. Выявляют5 ся пирамидные и экстрапирамидные симптомы: акинезия или  с псевдоспонтанными движениями, мио5 клония и повторяющиеся движения, не5 произвольное схватывание, постураль5 ная ретенция, рефлексы орального ав5 томатизма. К смертельному исходу мо5 жет привести вегетативный кризис с по5 вышением кровяного давления, тахи5
с псевдоспонтанными движениями, мио5 клония и повторяющиеся движения, не5 произвольное схватывание, постураль5 ная ретенция, рефлексы орального ав5 томатизма. К смертельному исходу мо5 жет привести вегетативный кризис с по5 вышением кровяного давления, тахи5
Кровоснабжение ствола мозга |
179 |
кардией, дыхательными нарушениями, гипертермией и профузным пото — и слюноотделением.
Если больной переживет эту фазу, за ней может следовать улучшение его состояния с постепенным возвратом контакта с окружающим. Однако после тотального поражения мозга обычно ос5 тается резидуальный психоорганиче5 ский синдром в сочетании с различ5 ными локальными неврологическими симптомами. Наблюдаются нарушения кратковременной и долговременной па5 мяти, ослабление концентрации внима5 ния, утрата инициативности, измене5 ния личности и регресс интеллекта. Иногда возможно удивительное улуч5 шение, особенно у больных молодого возраста.
Одной из причин описанного апал5 лического синдрома, по5видимому, яв5 ляется морфологическое и функцио5 нальное поражение ретикулярной фор5 мации среднего мозга. Кроме того, как следствие предшествующего отека, мо5 жет развиться генерализованное пора5 жение белого вещества полушарий го5 ловного мозга. После преходящей ос5 тановки сердца имеется вероятность по5 вреждения церебральной коры по обеим сторонам пограничных артериальных зон, максимально выраженного в те5 менно5затылочных областях, потому что они наиболее удалены от сердца.
13

4 Мозжечок
Внешнее строение
Мозжечок (cerebellum) и ствол мозга занимают заднюю черепную ямку, над которой в виде крыши расположен на5 мет мозжечка 
 от5 деляющий мозжечок от большого мозга (рис. 853). Каждая часть ствола мозга соединена с мозжечком парой ножек. Верхние мозжечковые ножки
от5 деляющий мозжечок от большого мозга (рис. 853). Каждая часть ствола мозга соединена с мозжечком парой ножек. Верхние мозжечковые ножки 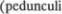 cerebellares
cerebellares  или соедини5 тельные плечи расположены на уровне среднего мозга; средние мозжечковые ножки (pedunculi cerebellares
или соедини5 тельные плечи расположены на уровне среднего мозга; средние мозжечковые ножки (pedunculi cerebellares 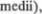 или плечи моста расположены на уровне моста и нижние мозжечковые ножки (pedunculi cerebellares
или плечи моста расположены на уровне моста и нижние мозжечковые ножки (pedunculi cerebellares  или ве5 ревчатые тела, расположены на уровне продолговатого мозга.
или ве5 ревчатые тела, расположены на уровне продолговатого мозга.
Наружная поверхность мозжечка, в отличие от поверхности больших по5 лушарий, имеет волнообразный вид, по5 скольку все извилины мозжечка — од5 ного малого диаметра и расположены параллельно друг другу подобно лист5 кам, разделенным узкими бороздками (рис. 4.1). Многочисленные ряды этих листков объединяются в различные бо5 лее крупные структуры, которые отде5 лены друг от друга более глубокими щелями. Полушария мозжечка связаны одной кортикализованной структурой, которая из5за своего внешнего вида получила название червь 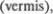 на сагиттальном разрезе она имеет почти циркулярную форму. Ее протяженность
на сагиттальном разрезе она имеет почти циркулярную форму. Ее протяженность
прерывается лишь вентрально, четвер5 тым желудочком. Ростральный конец червя называется язычком мозжечка  cerebelli), а ограничивающий же5 лудочек каудальный конец называется узелком (nodulus) (рис. 4.2). Кора узелка распространяется билатерально на об5
cerebelli), а ограничивающий же5 лудочек каудальный конец называется узелком (nodulus) (рис. 4.2). Кора узелка распространяется билатерально на об5  известные как
известные как 
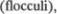 расположенные каудолате5 рально от средних и нижних мозжеч5 ковых ножек и отделенные от них бо5 ковыми карманами четвертого желудоч5 ка. Эти карманы связаны с субарахно5 идальным пространством в мосто5моз5 жечковом углу через отверстия
расположенные каудолате5 рально от средних и нижних мозжеч5 ковых ножек и отделенные от них бо5 ковыми карманами четвертого желудоч5 ка. Эти карманы связаны с субарахно5 идальным пространством в мосто5моз5 жечковом углу через отверстия  юшка (aperturae laterales
юшка (aperturae laterales  quarti). В об5 ласти отверстия может быть заметным выпячивающееся из него наподобие ма5 ленького букетика сосудистое сплетение
quarti). В об5 ласти отверстия может быть заметным выпячивающееся из него наподобие ма5 ленького букетика сосудистое сплетение  корзина
корзина  Ранее анатомы выделяли в полушариях и чер5 ве мозжечка множество частей, давая им различные названия. Такое деление коры мозжечка не имеет большого фун5 кционального или клинического значе5 ния. Наиболее важным оказалось выде5 ление в филогенетическом плане трех основных частей мозжечка:
Ранее анатомы выделяли в полушариях и чер5 ве мозжечка множество частей, давая им различные названия. Такое деление коры мозжечка не имеет большого фун5 кционального или клинического значе5 ния. Наиболее важным оказалось выде5 ление в филогенетическом плане трех основных частей мозжечка:
1.

2.
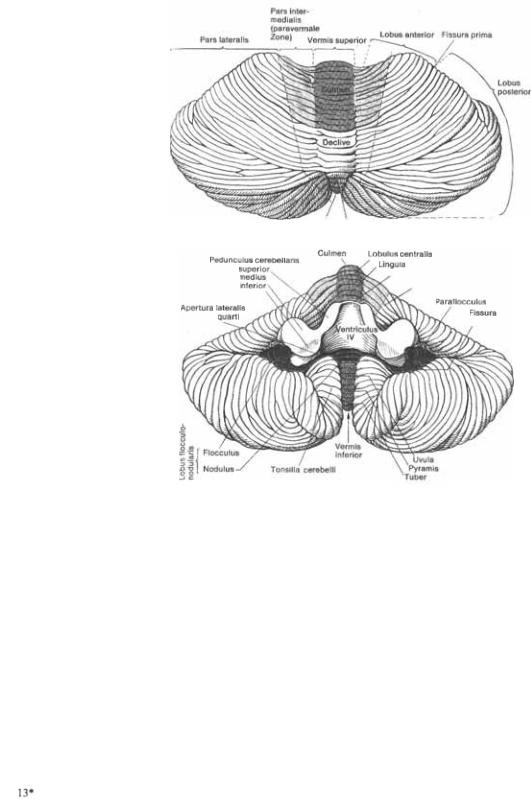
Внешнее строение |
181 |
 4.1Мозжечок(верх* няя поверхность).
4.1Мозжечок(верх* няя поверхность).
Tuber Folium
Velum 
Lobusanterior
ventriculi  lateralis
lateralis
 4.2Мозжечок(ниж* няя поверхность).
4.2Мозжечок(ниж* няя поверхность).
включает расположенную ростраль5 нее основной щели переднюю долю, вместе с язычком мозжечка, цент5 ральную дольку, вершину горки чер5 вя  с прилегающими сбоку зонами полушарий мозжечка; пира5 миду червя (pyramis vermis) и язычок нижнего червя (uvula vermis) вместе с околоклочковыми образованиями и миндалинами мозжечка (tonsillae
с прилегающими сбоку зонами полушарий мозжечка; пира5 миду червя (pyramis vermis) и язычок нижнего червя (uvula vermis) вместе с околоклочковыми образованиями и миндалинами мозжечка (tonsillae  Палеоцеребеллум представ5 ляет древнюю часть мозжечка. Он получает афферентные импульсы преимущественно по спинно5моз5 жечковым путям.
Палеоцеребеллум представ5 ляет древнюю часть мозжечка. Он получает афферентные импульсы преимущественно по спинно5моз5 жечковым путям.
3.  (neocerebellum), из5 вестный также как задняя доля, включает все части червя и полуша5 рий мозжечка, расположенные между основной и заднебоковой щелями. Это наиболее обширная и в филоге5 нетическом отношении наиболее мо5 лодая часть мозжечка. Ее развитие тесно связано с развитием коры больших полушарий и выпрямлени5 ем позы приматов. Точные движе5 ния, порождаемые двигательной ко5 рой больших полушарий, модифи5 цируются неоцеребеллумом. Анало5 гично развитию неокортикальных
(neocerebellum), из5 вестный также как задняя доля, включает все части червя и полуша5 рий мозжечка, расположенные между основной и заднебоковой щелями. Это наиболее обширная и в филоге5 нетическом отношении наиболее мо5 лодая часть мозжечка. Ее развитие тесно связано с развитием коры больших полушарий и выпрямлени5 ем позы приматов. Точные движе5 ния, порождаемые двигательной ко5 рой больших полушарий, модифи5 цируются неоцеребеллумом. Анало5 гично развитию неокортикальных
182 4 Мозжечок
Клетка |
Stratum |
|
culare |
||
|
Corpus 
|
dentatus |
волокна |
Мшистое волокно |
|
 4.3
4.3 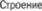 коры мозжечкасаффе* рентными и эфферентными связями.
коры мозжечкасаффе* рентными и эфферентными связями.
структур большого мозга, неоцере5 беллум 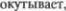 покрывает снару5 жи более древние части мозжечка.
покрывает снару5 жи более древние части мозжечка.
Эти три части мозжечка с учетом ос5 новных источников их афферентации могут быть обозначены как 
и 
Внутреннее строение
\
Перед обсуждением вопросов анатоми5 ческих и функциональных связей моз5 жечка с другими отделами центральной нервной системы необходимо остано5 виться на внутреннем строении моз5 жечка.
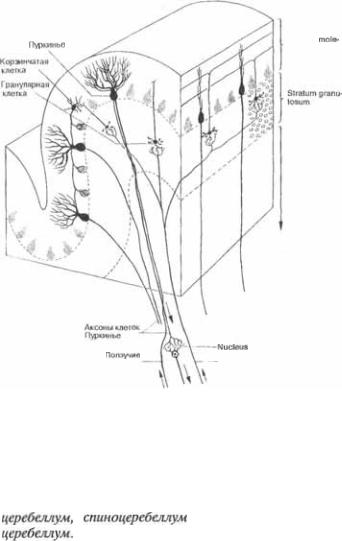
Кора мозжечка состоит всего из трех слоев — молекулярного слоя, слоя  ток Пуркинье и гранулярного слоя (рис. 4.3)
ток Пуркинье и гранулярного слоя (рис. 4.3)
Молекулярный слой содержит раз5  бросанные мелкие нейроны (звездчатые
бросанные мелкие нейроны (звездчатые  и корзинчатые клетки); разветвления дендритов нескольких типов
и корзинчатые клетки); разветвления дендритов нескольких типов 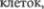 в
в  основном клеток Пуркинье; многочис5 ленные тонкие аксоны, преимуществен5 но от очень мелких гранулярных клеток гранулярного слоя. Слой клеток Пур5 кинье содержит только тела этих до5 вольно крупных клеток, расположенных бок о бок в один ряд. В противополож5 ность этому, гранулярные клетки гра5 нулярного слоя по своим размерам не превосходят лимфоциты и расположе5 ны довольно беспорядочно. Рассеянные между ними отдельные более крупные нейроны относятся к клеткам второго типа Гольджи. Немиелинизированные аксоны гранулярных
основном клеток Пуркинье; многочис5 ленные тонкие аксоны, преимуществен5 но от очень мелких гранулярных клеток гранулярного слоя. Слой клеток Пур5 кинье содержит только тела этих до5 вольно крупных клеток, расположенных бок о бок в один ряд. В противополож5 ность этому, гранулярные клетки гра5 нулярного слоя по своим размерам не превосходят лимфоциты и расположе5 ны довольно беспорядочно. Рассеянные между ними отдельные более крупные нейроны относятся к клеткам второго типа Гольджи. Немиелинизированные аксоны гранулярных 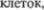 поднима5 ющиеся в молекулярный слой, распо5 ложены в нем только параллельно про5
поднима5 ющиеся в молекулярный слой, распо5 ложены в нем только параллельно про5
дольной оси каждой 
 листка. Напротив, дендриты клеток
листка. Напротив, дендриты клеток
Пуркинье и корзинчатых клеток рас5 пространяются в плоскостях перпенди5 кулярно продольной оси каждого лист5 ка, то есть параллельно червю (рис. 4.3).
Клетки Пуркинье — единственные нейроны мозжечка, передающие эффе5 рентные импульсы из коры мозжечка. Их аксоны, начинаясь от тела клетки, идут через гранулярный слой и белое вещество к глубоким мозжечковым яд5 рам. Их дендриты получают всеобъем5 лющую информацию от различных от5 делов центральной нервной системы, как прямым, так и непрямым путем, через релейные пункты. Часть импуль5 сов — стимулирующие, часть — инги5 бирующие. Указанная информация по5 ступает по отросткам различных ней5 ронов, именуемым как «карабкающие5 ся»,  «корзинчатые», «танге5 циальные» и «параллельные»волокна. Мшистые волокна передают импульсы почти исключительно из спинного моз5 га, вестибулярных ядер и ядер моста; в качестве передатчиков информации служат гранулярные клетки грануляр5 ного слоя. Карабкающиеся волокна пе5 редают импульсы из ядер нижних олив непосредственно на дендриты клеток Пуркинье.
«корзинчатые», «танге5 циальные» и «параллельные»волокна. Мшистые волокна передают импульсы почти исключительно из спинного моз5 га, вестибулярных ядер и ядер моста; в качестве передатчиков информации служат гранулярные клетки грануляр5 ного слоя. Карабкающиеся волокна пе5 редают импульсы из ядер нижних олив непосредственно на дендриты клеток Пуркинье.
Каждое полушарие мозжечка имеет четыре парных ядра: ядро шатра, ша5 ровидное, пробковидное и зубчатое (nucleus  nucleus globosus, nucleus
nucleus globosus, nucleus  nucleus dentatus). На
nucleus dentatus). На  3.48 эти ядра изображены на попереч5 ном разрезе через мост и белое вещество мозжечка. Рис. 4.4 показывает их рас5 положение на сагиттальном разрезе мозжечка. Первые три ядра находятся в крыше четвертого желудочка. Ядро шатра локализуется вблизи конька шат5 ра
3.48 эти ядра изображены на попереч5 ном разрезе через мост и белое вещество мозжечка. Рис. 4.4 показывает их рас5 положение на сагиттальном разрезе мозжечка. Первые три ядра находятся в крыше четвертого желудочка. Ядро шатра локализуется вблизи конька шат5 ра  Оно наиболее древнее в филогенетическом отношении и полу5 чает афферентацию от архицеребеллу5 ма, точнее, от флоккулонодулярной зо5
Оно наиболее древнее в филогенетическом отношении и полу5 чает афферентацию от архицеребеллу5 ма, точнее, от флоккулонодулярной зо5
Внутреннее строение |
183 |
ны, и частично от червя. Его эфферент5 ные волокна направляются через ниж5 нюю мозжечковую ножку к вестибуляр5 ным ядрам (шатрово5бульбарный, или мозжечково5бульбарный тракт). Много5 численные волокна переходят на другую сторону мозжечка, делая петлю вокруг контрлатеральной верхней мозжечковой ножки и достигая ретикулярной фор5 мации и вестибулярных ядер посредст5 вом дугообразного пучка Рассела. Ша5 ровидное и пробковидное ядра залегают несколько латеральнее ядра шатра. Они получают афферентацию от соседней с червем области палеоцеребеллума. Их эфферентные пути идут к контрлате5 ральным красным ядрам через верхние мозжечковые ножки (рис. 4.4). Зубчатое ядро5самое крупное  всех четырех ядер, оно расположено в центральной части белого вещества полушария моз5 жечка. Оно получает импульсы от кле5 ток Пуркинье коры всего неоцеребел5 лума и части палеоцеребеллума. Эффе5 рентные волокна идут в верхних ножках мозжечка, переходят на противополож5 ную сторону на границе моста и сред5 него мозга, оканчиваются в контрлате5 ральном красном ядре и вентролате5 ральном ядре таламуса (рис. 55). От таламуса волокна идут к первичной дви5 гательной коре (поля Бродмана 4 и 6).
всех четырех ядер, оно расположено в центральной части белого вещества полушария моз5 жечка. Оно получает импульсы от кле5 ток Пуркинье коры всего неоцеребел5 лума и части палеоцеребеллума. Эффе5 рентные волокна идут в верхних ножках мозжечка, переходят на противополож5 ную сторону на границе моста и сред5 него мозга, оканчиваются в контрлате5 ральном красном ядре и вентролате5 ральном ядре таламуса (рис. 55). От таламуса волокна идут к первичной дви5 гательной коре (поля Бродмана 4 и 6).
Все поступающие в мозжечок им5 пульсы оканчиваются в коре мозжечка или, через коллатерали, в ядрах моз5 жечка. Эти афференные импульсы на5 чинаются в коре больших полушарий, в стволе (вестибулярных ядрах, ретику5 лярной формации, нижней оливе, до5 бавочном клиновидном ядре) и в спин5 ном мозге. Через нижние ножки моз5 жечка (веревчатые тела) проходят сле5 дующие афферентные волокна:
1.Волокна от вестибулярных ядер, окан5 чивающиеся в клочково5узелковой доле (связанной с ядром шатра).
2.Оливомозжечковый путь (tr. olivocere5 bellaris), начинающийся в контрлате5
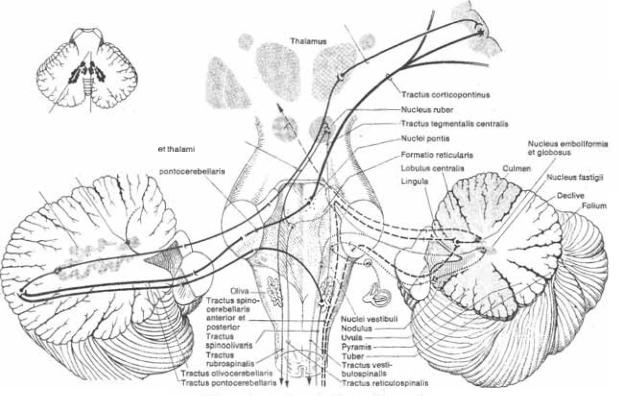
Tractus 
Плоскости сечений
Tractus 
Tractus
Nucleus 
Nucleus 
 4.4 Афферентные и эфферентные проводящие пути мозжечка (слева * через зубчатое ядро; справа * через червь). На вставке показаны плоскости разрезов.
4.4 Афферентные и эфферентные проводящие пути мозжечка (слева * через зубчатое ядро; справа * через червь). На вставке показаны плоскости разрезов.
ральной нижней оливе и оканчива5 ющийся в виде «карабкающихся» во5 локон на дендритах клеток Пуркинье мозжечка.
Задний спинно5мозжечковый путь
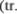
 posterior), кото5 рый начинается от столба Кларка (грудного ядра, nucleus thoracicus), расположенного у основания заднего рога спинного мозга (рис. 1.19). Этот путь проводит импульсы из мышеч5 ных веретен в находящиеся рядом с червем зоны передней и задней долей мозжечка. Указанный путь состоит из наиболее быстро5проводящих во5 локон во всей нервной системе.
posterior), кото5 рый начинается от столба Кларка (грудного ядра, nucleus thoracicus), расположенного у основания заднего рога спинного мозга (рис. 1.19). Этот путь проводит импульсы из мышеч5 ных веретен в находящиеся рядом с червем зоны передней и задней долей мозжечка. Указанный путь состоит из наиболее быстро5проводящих во5 локон во всей нервной системе.
4. Волокна, начинающиеся в добавоч5 ном клиновидном ядре и присоеди5 няющиеся к заднему спинно5моз5 жечковому пути. Эти волокна прово5 дят импульсы, поступающие в доба5 вочное клиновидное ядро из ядер средней и оральной частей шейного отдела спинного мозга, находящихся выше уровня столба Кларка; они про5
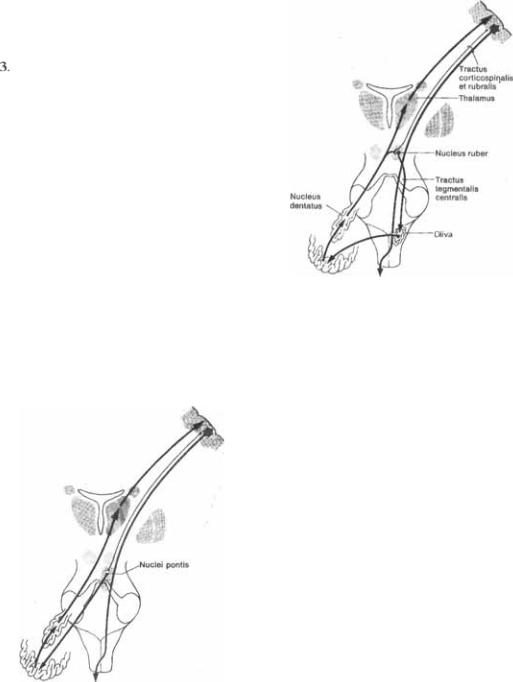
 4.5 Мозжечковая цепь обратной связи через ядра
4.5 Мозжечковая цепь обратной связи через ядра  моста.
моста.
Внутреннее строение |
185 |
Рис. 4.6 Мозжечковая  обратной связи через ниж* нюю оливу с вовлечением треугольника Гюллейн*Мол* ларета (красное ядро, центральный покрышечный путь, олива, мозжечок, красное ядро).
обратной связи через ниж* нюю оливу с вовлечением треугольника Гюллейн*Мол* ларета (красное ядро, центральный покрышечный путь, олива, мозжечок, красное ядро).
ходят в латеральной части клиновид5 ного пучка.
5.Волокна от ретикулярной формации ствола мозга (на рис. 4.4 не показа5
ны).
По нижней ножке мозжечка проходит один эфферентный путь — шатрово5 бульбарный, или мозжечково5бульбар5 ный тракт, направляющийся к вести5 булярным ядрам и представляющий эфферентное звено вестибуломозжечко5 вого модулирующего кольца обратной связи. Посредством него  влияет на двигательную активность спинного мозга через вестибуло5спин5 номозговой путь и медиальный про5 дольный пучок.
влияет на двигательную активность спинного мозга через вестибуло5спин5 номозговой путь и медиальный про5 дольный пучок.
Средняя ножка  (плечо моста) состоит в основном из перекре5 щивающихся мосто5мозжечковых воло5 кон. Они идут от нейронов ядер моста, являющихся вторыми нейронами кор5 ково5ядерных путей.
(плечо моста) состоит в основном из перекре5 щивающихся мосто5мозжечковых воло5 кон. Они идут от нейронов ядер моста, являющихся вторыми нейронами кор5 ково5ядерных путей.
Верхняя ножка мозжечка (соедини5
186 4 Мозжечок
тельное плечо) содержит эфферентные волокна, начинающиеся от нейронов зубчатых, шаровидных, пробковидных ядер и ядер шатра. Эти волокна следуют к контрлатеральному красному ядру, вентролатеральному и срединно5меди5 альному ядрам таламуса, ретикулярной формации ствола.
Постсинаптические  вые волокна идут к коре, от которой нисходят корковомостовые волокна. Та5 ким образом замыкается важное кольцо обратной связи, распространяющееся от коры больших полушарий к ядрам мос5 та, от
вые волокна идут к коре, от которой нисходят корковомостовые волокна. Та5 ким образом замыкается важное кольцо обратной связи, распространяющееся от коры больших полушарий к ядрам мос5 та, от  к коре мозжечка, к зубчатому ядру и далее вновь к таламусу и коре (рис. 45; 4.6).
к коре мозжечка, к зубчатому ядру и далее вновь к таламусу и коре (рис. 45; 4.6).
Дополнительная цепь обратной свя5 зи образована треугольником Гюллейн5 Молларета — от красного ядра к ниж5 ней оливе через центральный покры5 шечный тракт, оттуда к мозжечковой коре, к зубчатому ядру и вновь к крас5 ному ядру (рис. 4.6). Таким образом, мозжечок опосредованно модулирует двигательную активность спинного мозга через свои связи с красными яд5 рами и ретикулярной формацией, от которых начинаются нисходящие красноядерно5спинномозговые и рети5 кулярно5спинномозговые тракты. Эф5 фект мозжечковых влияний ипсилате5 ральный, что обусловлено двойным пе5 рекрестом путей в этой системе. Волок5 на, идущие от 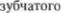 ядра, пересе5 каются и достигают противоположного красного ядра. Волокна нисходящего красноядерно5спинномозгового тракта сразу же после выхода из красного ядра вновь переходят на противоположную сторону — перекрест Фореля.
ядра, пересе5 каются и достигают противоположного красного ядра. Волокна нисходящего красноядерно5спинномозгового тракта сразу же после выхода из красного ядра вновь переходят на противоположную сторону — перекрест Фореля.
Верхняя мозжечковая ножка содер5 жит лишь один афферентный путь — передний спинно5мозжечковый тракт (tr. spinocerebellaris anterior), который оканчивается в палеоцеребеллуме, как и задний спинно5мозжечковый тракт (tr. spinocerebellaris posterior). Оба спин5
но5мозжечковых тракта передают про5 приоцептивные импульсы от перифе5 рических рецепторов — мышечных ве5 ретен, сухожильных органов Гольджи и рецепторов, воспринимающих давле5 ние. Импульсы от мышечных веретен проводятся в основном по задним  но5мозжечковым путям, а импульсы от сухожильных рецепторов Гольджи — преимущественно по передним спинно5 мозжечковым путям.
но5мозжечковым путям, а импульсы от сухожильных рецепторов Гольджи — преимущественно по передним спинно5 мозжечковым путям.
Таким образом, мозжечок получает чувствительную информацию от всех отделов центральной нервной системы по волокнам, проходящим в составе трех ножек. Он также связан со всеми двигательными путями, поскольку его ядра являются источниками эфферент5 ной импульсации в регулирующих кру5
 обратной связи. Хотя мозжечок свя5 зан с корой больших полушарий через
обратной связи. Хотя мозжечок свя5 зан с корой больших полушарий через 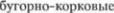 пути, его деятель5 ность не контролируется сознанием. В
пути, его деятель5 ность не контролируется сознанием. В
этом заключается одна из причин слож5 ности определения нормальной функ5 ции мозжечка. Все, что известно об этой структуре, получено при обобщении данных эмбриологии, сравнительной анатомии, результатов экспериментов и клинических описаний случаев с точ5 но известной локализацией очага по5 ражения в определенных отделах моз5 жечка.
Функция
Мозжечок, являясь частью комплексно5 го регуляторного механизма с обратной связью, действует в качестве координа5 ционного центра по обеспечению рав5 новесия тела и поддержанию мышеч5 ного тонуса. Он также обеспечивает воз5 можность выполнения дискретных и точных движений. Предполагается, что мозжечок работает подобно компьюте5 ру, отслеживая и координируя сенсор5 ную информацию на входе и модулируя моторные сигналы на выходе.
Архицеребеллум получает информа5 цию о положении головы в пространст5 ве от вестибулярной системы и о дви5 жениях головы — от рецепторов полу5 кружных каналов посредством кинесте5 тических импульсов. Это позволяет мозжечку постоянно синергично мо5 дулировать двигательные импульсы спинного мозга, что обеспечивает со5 хранение равновесия вне зависимости от положения тела и его движения.
Поражение клочково5узелковой до5 ли вызывает нарушение равновесия и устойчивости при стоянии (астазия) и ходьбе (абазия). Наблюдается шаткая походка с широко расставляемыми но5 гами, похожая на походку пьяного (атаксия туловища или осевая атаксия). Эта атаксия не увеличивается при за5 крывании глаз в противоположность атаксии, вызванной поражением задних канатиков спинного мозга. Мозжечко5 вая атаксия связана не с патологиче5 ским сниженим проприоцептивной чув5 ствительности, а с нарушением возмож5 ности осуществления координирован5 ной работы мышц, называемой  гией (клочково5узелковый синдром).
гией (клочково5узелковый синдром).
Поражение узелка (nodulus) наруша5 ет реакцию на калорическую и враща5 тельную пробы, применяемые при ис5 следовании вестибулярного анализато5 ра. Пациенты, у которых были удалена часть узелка и язычка червя (uvulae), не чувствительны к укачиванию. Такая же утрата функции возникает при пре5 рывании афферентных или эфферент5 ных волокон клочково5узелковой  Равновесие поддерживается следующей рефлекторной дугой: импульсы из ла5 биринта поступают прямым путем либо опосредованно в вестибулярные ядра, от них — в архицеребеллум и в ядра шатра, откуда эфферентные импульсы возвращаются в латеральные вестибу5 лярные ядра
Равновесие поддерживается следующей рефлекторной дугой: импульсы из ла5 биринта поступают прямым путем либо опосредованно в вестибулярные ядра, от них — в архицеребеллум и в ядра шатра, откуда эфферентные импульсы возвращаются в латеральные вестибу5 лярные ядра 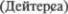 и ретикуляр5 ную формацию.
и ретикуляр5 ную формацию.  номозговому, ретикулярно5спинномоз5
номозговому, ретикулярно5спинномоз5
Функция 187
говому путям и медиальному продоль5 ному пучку импульсы достигают клеток передних рогов спинного мозга и мо5 дулируют их активность. Повреждение этой системы может вызвать нистагм; других мозжечковых симптомов, кроме описанных выше, не наблюдается.
Палеоцеребеллум получает аффе5 рентные импульсы из спинного мозга по переднему и заднему спинно5моз5 жечковым трактам и от добавочного клиновидного ядра по клиновидно5моз5 жечковому пути. Эфферентные импуль5 сы из палеоцеребеллума модулируют активность антигравитационной муску5 латуры и обеспечивают мышечный то5 нус, необходимый для поддержания равновесия при стоянии или ходьбе. Спинномозговые импульсы имеют со5 матотопическую проекцию на кору па5 леоцеребеллума, в каждом полушарии мозжечка представлена ипсилатераль5 ная часть тела. Кора прилежащих к чер5 вю областей проецируется на шаровид5 ное и пробковидное ядра, кора червя — на ядра шатра.
Эфферентные волокна от нейронов глубоких ядер мозжечка пересекаются в верхних мозжечковых ножках и до5 стигают контрлатеральных красных ядер. Нисходящие красноядерно5спин5 номозговые и красноядерно5ретикуляр5 ные пути вновь пересекаются и моду5 лируют активность двигательных ней5 ронов ствола мозга и спинного мозга, расположенных ипсилатерально по от5 ношению к ядрам мозжечка и контр5 латералыю по отношению к красным ядрам. Импульсы от мозжечковых ядер также проецируются на центрально5сре5 динные ядра таламуса и от них на хво5 статые ядра и скорлупу (стриатум), влияя таким образом на  мидную систему. Содружественное дей5 ствие палеоцеребеллума и архицеребел5 лума обеспечивает регуляцию мышеч5 ного тонуса и тонкую координирован5 ную работу мышц агонистов и антаго5
мидную систему. Содружественное дей5 ствие палеоцеребеллума и архицеребел5 лума обеспечивает регуляцию мышеч5 ного тонуса и тонкую координирован5 ную работу мышц агонистов и антаго5