

Механизмы получения энергии
Часть свободной энергии, высвобождаемой в окислительно-восстановительных реакциях, может быть использована для синтеза АТФ, который служит важнейшим, но не единственным переносчиком энергии в клетке. Помимо АТФ в метаболизме клетки участвуют другие богатые энергией соединения, к ним относятся другие нуклеотид ди- и трифосфаты, аденозинфосфосульфат, ацилфосфаты (например, карбамоилфосфат), фосфоенолпируват. * Важнейшим макроэргическим соединением для анаэробов служит ацетил-КоА. Однако все эти соединения обмениваются с АТФ и поэтому представления об АТФ, как универсальном переносчике энергии, достаточно.
Показаны области развития основных групп железобактерий. Пунктир — попе устойчивости воды
АТФ может синтезироваться путем субстратного фосфори-лирования, чаще всего за счет переноса фосфорильной группы от богатого энергией соединения на АДФ. Такой способ реализуется при гликолизе, разнообразных видах брожения и некоторых других процессах. Субстратное фосфорилирование осуществляется в цитоплазме и может быть воспроизведено в бесклеточных экстрактах.
Второй механизм синтеза АТФ, мембранное фосфорилирование, необходимо зависит от наличия энергизованной мембраны и связан с использованием энергии трансмембранного электрохимического градиента ионов (в общем случае Дц1), чаще всего ионов водорода (ДцН). Этот механизм реализуется в процессах дыхания (окислительное фосфорилирование) и фотосинтеза (фо-тофосфорилирование).
Окислительно-восстановительные реакции, в результате которых бактерии получают энергию, представляют собой перенос электронов от окисляемого вещества (донора электронов) к окислителю (акцептору). Перенос электронов от донора к акцептору осуществляется по градиенту редокс-потенциала через ряд последовательно функционирующих переносчиков. Часть из них закреплена в мембране, а часть находится в цитоплазме и сопрягает эти два компонента клетки. Набор переносчиков характерен для каждого организма.
Комплекс переносчиков электронов работает как универсальная структура для всех окисляемых веществ, лишь бы в метаболизме была реакция, совмещающая окислительное преобразование субстрата с цепью переноса электронов, на конце которой находится окислитель, например для О2 — цитохромоксидаза. Вместе с тем универсальным продуктом реакции должна быть генерация АТФ. Она осуществляется мембранным ферментом АТФ-синтазой, катализирующим синтез АТФ путем конверсии энергии трансмембранного электрохимического градиента протонов цН в энергию АТФ.
У аэробных органотрофов донором электронов служат органические вещества, окисление которых приводит к восстановлению переносчика водорода НАД в НАДН. Последний реагирует с электронтранспортной цепью. Моделью для окислительного фосфорилирования служит митохондрия эукариот.
Суммарный процесс переноса электронов разбит на три стадии, которые катализируются комплексами I, III и IV. Каждый комплекс представляет собой весьма крупное образование, построенное из многих полипептидных цепей. Первая стадия окисления НАДН катализируется комплексом I (НАДН: убихинон-оксидоредуктазой) и ведет к восстановлению убихинона Q. В состав комплекса I входит флавопротеин и FeS-кластеры. Вторая стадия, реакция окисления восстановленного убихинона и сопряженного восстановления цит с катализируется комплексом III (убихинон : цит с-оксидоредуктазой или всцитохромным комплексом). В его состав входит FeS-белок Риске, цит с, или /, цит в. Третья стадия, перенос электронов от цит с на Ог, катализируется комплексом IV, цитохромоксидазой. Перенос электронов сопряжен с переносом протонов через сопрягающую мембрану и генерацией ДцН. Превращение энергии ДцН в энергию АТФ осуществляется АТФ-синтазой.
У фотосинтезирующих организмов процессы преобразования световой энергии, приводящие к синтезу АТФ и образованию восстановителя, необходимого, например, для фиксации СО2, включают следующие этапы. Энергия света поглощается пигментами светособирающих антенных комплексов и передается в реакционный центр (РЦ), в котором происходит превращение электромагнитной формы энергии в энергию электрохимического разделения разрядов. Фотохимически активный хлорофилл (или бактериохлорофилл) РЦ, обозначаемый обычно буквой Р (от англ. pigment), переходит в электронно-возбужденное состояние и окисляется, отдавая электрон первичному акцептору. Перенос электрона на вторичный акцептор и восстановление окисленного Р первичным донором электронов стабилизирует разделение зарядов в РЦ. Дальнейшие процессы связаны с переносом электрона по электронтранспортнои цепи, сопряженным с синтезом АТФ
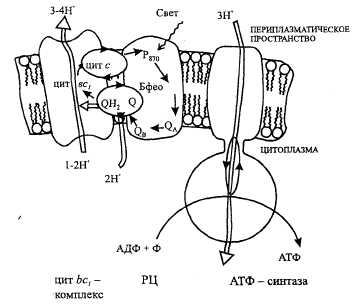
Рис. 5. Цепь переноса электронов при фотосинтезе А. Аноксигенный фотосинтез (пурпурные бактерии)
Р870 — бактериохлорофилп; Бфео — бактериофеофитин; Q — хинон (Q, — плас-тохинон); цит — цитохром; Р6в0, Р7Ю — хлорофилл; Фео — феофитин; Фд — фер-редоксин; ПЦ — пластоцианин; ФС — фотосистема; РЦ — реакционный центр;
ФНР — ферредоксии:НАДФ оксидоредуктаза.
Электроны от фотовозбужденного Рв70 передаются на убихинон Q, восстанавливая его, далее на цит Ьсу комплекс и через цит сие, возвращаются к молекуле окисленного Р870. Перенос электронов сопряжен с трансмембранным переносом протонов при окислении убихинола цит be, комплексом, что ведет к генерированию ДцН. Результатом циклического переноса электронов является синтез АТФ.
Б. Оксигенный фотосинтез
У оксигенных фототрофов имеются две фотосистемы: ФС I (с Р700) и ФС II (с Р680). В состав ФС II входит Mn-содержащий комплекс, осуществляющий разложение Н2О. Электроны от фотовозбужденного Р680 через ряд переносчиков РЦ передаются на пластохинон, палее в цит Ье1 комплекс и через пластоцианин на Р700. От фотовозбужденного Р700 (через ряд переносчиков РЦ) электроны поступают на Фд, связывающий ФС I с НАДФ-редуктазой (ФНР), которая восстанавливает НАДФ. Таким образом, происходит последовательный перенос электронов от воды к НАДФ. Реакции электронного транспорта, связанные с разложением воды в ФС II и окислением молекулы лластохинола цит комплексом, сопровождаются трансмембранным переносом протонов, сопряженным с генерированием ДцН и синтезом АТФ. Образование АТФ возможно и в результате связанного с ФС I циклического преноса электронов (показан пунктиром).
Природа акцепторов электронов в РЦ и состав переносчиков электронов у разных групп фототрофов различаются. Существенно, что часть переносчиков являются общими для фотосинтетической и дыхательной электронтранспортнои цепи, обусловливая у бактерий взаимосвязь процессов фотосинтеза и дыхания.
Различают циклический или нециклический пути переноса электронов. Циклический путь, свойственный, например, пурпурным бактериям, обеспечивает синтез АТФ (см. рис. 5 А), но не восстановителя. Поэтому восстановление НАД у этих микроорганизмов происходит в результате АТФ-зависимого обратного переноса электронов. У зеленых серобактерий перенос электронов осуществляется по нециклическому пути, обеспечивая синтез АТФ и восстановление НАД. Донорами электронов у аноксиген-ных, т. е. не выделяющих О2, организмов служат восстановленные соединения серы, S°, H2, Fe2+ или некоторые органические вещества. Цианобактерии и эукариотные фототрофы используют в качестве донора электронов Н2О и осуществляют фотосинтез с выделением О2 — оксигенный. В отличие от аноксигенных бактерий, имеющих одну фотосистему (ФС), у них последовательно функционируют две фотосистемы (см. рис. 5 Б) и может осуществляться как нециклический (так называемая Z-схема), так и индуцируемый ФС I циклический перенос электронов. Окисление Н2О осуществляется Mn-содержащим комплексом, связанным с ФС П. Особый тип фотосинтеза известен у экстремально галофильных архебактерий (галобактерий). В этом случае трансмембранный перенос Н+ и конверсия энергии света в энергию А\хН с последующим синтезом АТФ осуществляются в результате светоиндуцированных преобразований бактериородопсина — пигмент-белкового комплекса, содержащего каротиноид ретиналь.
Окислительное фосфорилирование. Сопряжение окисления и фосфорилирования в митохондриях довольно прочно. Фосфорилирование сопряжено с переносом электронов и поэтому называется окислительным фосфорилированием. Если невозможен синтез АТФ, то прекращается перенос электронов в дыхательной цепи.
При переносе электронов по дыхательной цепи происходит выделение энергии. На некоторых участках ЦПЭ выделившейся энергии достаточно для образования макроэргической связи в молекуле АТФ. Это участки сопряжения окисления и фосфорилирования, всего в ЦПЭ их три. Количество этих участков определено путем измерения коэффициента фосфорилирования. Коэффициент фосфорилирования – это отношение связанного фосфора к поглощенному кислороду – Р/О. Если субстрат окисляется при участии НАД-зависимой ДГ (яблочная, молочная, альфа-кетоглутаровая, изолимонная кислоты) образуется 3 АТФ, ФАД-зависимой (янтарная кислота, амины, ацил-КоА) – 2 АТФ. Б. Чанс, используя спектрометрический метод определил положение участков сопряжения окисления и фосфорилирования:
1) между НАД и ФАД
2) между цитохромом в и цитохромом с
3) на участке цитохромоксидазы.
На этих трех участках выделяется энергии больше, чем 34,5 кДж/моль, поэтому возможен синтез АТФ. Таким образом, дыхательная цепь разделяет энергию, выделяющуюся при переносе электронов, на отдельные «порции».
Как энергия электронов дыхательной цепи трансформируется в энергию фосфатных связей АТФ объясняет хемиоосмотическая теория Митчелла – Скулачева. Согласно этой теории, во внутренней мембране митохондрий имеются ферменты – протонные насосы, которые используя энергию электронов дыхательной цепи, выкачивают протоны из матрикса в межмембранное пространство. В результате этого процесса наружная часть мембраны получает положительный заряд. В матриксе митохондрий при этом образуется избыток гидроксильных ионов, внутренняя сторона мембраны заряжается отрицательно. Таким образом, на сопрягающей мембране возникает одновременно с градиентов концентрации ионов водорода градиент электрического потенциала. Обратный поток протонов по градиенту концентрации к матриксной стороне мембраны осуществляется через АТФ-синтетазу. АТФ-синтетаза – протонный насос, использующий движение протонов вниз по градиенту для того, чтобы синтезировать АТФ. Нобелевская премия 1997 года за прямую демонстрацию механохимических свойств Н+-АТФазной системы.
Определения понятия жизни и гипотезы происхождения жизни на Земле.
«Жизнь есть способ существования белковых тел, и этот способ существования состоит по своему существу в постоянном самообновлении химических составных частей этих тел» - Ф. Энгельс. Жизнь - это высшая по сравнению с физической и химической форма существования материи. Живые объекты отличаются от неживых обменом веществ - непрерывным условием жизни, способностью к размножению, росту, активной регуляции своего состава и функций, к различным формам движения, раздражимостью, приспособляемостью к среде и т. д. Окончательного и строгого определения понятия «жизнь» нет до сих пор. Неясно, можно ли вирусы считать формой жизни, т. к. вне клеток хозяина они не проявляют никаких признаков жизни, например, они не могут размножаться.
Живые системы характеризуются очень высоким уровнем структурной и функциональной организации на молекулярном уровне, высочайшей информационной плотностью, самоорганизацией, способностью к самовосстановлению и т. п.
Существуют следующие пять основных групп теорий о происхождении жизни.
1. Креационизм, утверждающий, что жизнь была создана сверхъестественным существом (Богом, космическим разумом и т.п.), к этому направлению примыкают теологи и философы-идеалисты. Этот процесс был произведен один раз, больше он не повторится и поэтому не доступен экспериментальной проверке. Поэтому эта теория выходит за рамки научного исследования.
Остальные направления материалистичны.
2. Теория самопроизвольного зарождения - жизнь самозарождается при создании для этого подходящих условий, и это на протяжении всей истории Земли на ней происходило неоднократно, однако попытки создания жизни в искусственных лабораторных условиях (в «пробирке») химическим путем пока не удались.
3. Теория стационарного состояния. Жизнь существовала всегда, и только изменялись ее формы.
4. Теория панспермии. Жизнь на Землю была занесена из космоса, поскольку в нем зародыши жизни и белковые элементы непрерывно переносятся с планеты на планету (подтверждено исследованиями).
5. Теория биохимической революции - жизнь произошла естественным путем в результате саморазвития химических и физических процессов (примыкает ко второй группе теорий).
Теория абиогенного происхождения жизни
Пожалуй, первая научная, хорошо продуманная теория происхождения жизни абиогенным путем была предложена биохимиком А.И. Опариным еще в 20-х годах прошлого века и дополненная Дж. Холдейном. Теория базировалась на представлении, что все начиналось с белков, и на возможности в определенных условиях спонтанного химического синтеза мономеров белков - аминокислот и белковоподобных полимеров (полипептидов) абиогенным путем. Публикация теории стимулировала многочисленные эксперименты в ряде лабораторий мира, показавшие реальность такого синтеза в искусственных условиях. Теория быстро стала общепринятой и необыкновенно популярной.
Основным ее постулатом было то, что спонтанно возникавшие в первичном «бульоне» белковоподобные соединения объединялись в коацерватные капли - обособленные коллоидные системы (золи), плавающие в более разбавленном водном растворе. Это давало главную предпосылку возникновения организмов - обособление некой биохимической системы от окружающей среды, ее компартментализацию. Так как некоторые белковоподобные соединения коацерватных капель могли обладать каталитической активностью, то появлялась возможность прохождения биохимических реакций синтеза внутри капель - возникало подобие ассимиляции, а значит, роста коацервата с последующим его распадом на части - размножением. Ассимилирующий, растущий и размножающийся делением коацерват рассматривался как прообраз живой клетки.
У теории Опарина-Холдейна имеются существенные недостатки – она не объясняла механизма возникновения из коацерватов полноценной клетки и способности к самовоспроизведению. Существует и ряд других проблем – каким образом аминокислоты без участия ферментов могли образовать белковые структуры, как образовались первые ферменты и т.д. Для объяснения этих моментов теория модифицировалась, и этот процесс продолжается до сегодняшнего дня.
По господствующим современным представлениям, образованию белков предшествовало образование РНК. Нуклеиновые кислоты являются единственным типом биологических полимеров, макромолекулярная структура которых, благодаря принципу комплементарности при синтезе новых цепей, обеспечивает возможность копирования собственной линейной последовательности мономерных звеньев, другими словами, возможность воспроизведения (репликации) полимера, его микроструктуры. Поэтому только нуклеиновые кислоты, но не белки, могут быть генетическим материалом, то есть воспроизводимыми молекулами, повторяющими свою специфическую микроструктуру в поколениях.
В настоящее время общепринято считать следующие эры развития жизни на Земле:
- Архейская эра (начало жизни; считается, что жизнь возникла 3,5 млрд. лет назад);
- Протерозойская эра (одноклеточные, беспозвоночные, водоросли);
- Палеозойская эра (растения (псилофит), кистеперые рыбы, земноводные, насекомые);
- Мезозойская эра (пресмыкающиеся, млекопитающие);
- Кайнозойская эра (млекопитающие, птицы, человек).
Развитие жизни на Земле
Геологическая и палеонтологическая летописи тесно взаимосвязаны. В осадочных толщах разного возраста встречаются ископаемые остатки животных и растений. Поэтому историю развития жизни на Земле делят на геологические эры и периоды.
По степени геологической и палеонтологической изученности история земной коры подразделяется на две неравные части. Более молодой интервал времени, охватывающий последние 570 млн. лет, относительно хорошо изучен. Американским геологом Ч. Шухертом он назван фанерозойским эоном или фанерозоем (от греч. «фанерос»— очевидный, четкий, «зое»— жизнь). К фанерозою относятся кайнозойская, мезозойская и палеозойская эры. Вторая, более древняя и продолжительная часть названа криптозойским эоном (период со скрытым развитием жизни). Он охватывает огромный интервал времени — 4600—570 млн. лет назад. В большинстве случаев организмы криптозоя не имели твёрдого скелета и оставили довольно скудные следы.
Криптозой. Архей - древнейшая геологическая эра Земли (3,5 - 2,6 млрд. лет назад).
Ко времени архея относится возникновение первых прокариот (бактерий и сине-зеленых) - организмов, которые в отличие от эукариот не обладают оформленным клеточным ядром и типичным хромосомным аппаратом (наследственная информация реализуется и передается через ДНК).
В отложениях архея найдены также остатки нитчатых водорослей. В этот период появляются гетеротрофные организмы не только в море, но и на суше. Образуется почва. В атмосфере снижается содержание метана, аммиака, водорода, начинается накопление углекислого газа и кислорода.
Протерозой (с греч. «первичная жизнь») - огромный по продолжительности этап исторического развития Земли (2,6 млрд.-570 млн. лет назад). Возникновение многоклеточности - важный ароморфоз в эволюции жизни. Конец протерозоя иногда называют «веком медуз» - очень распространенных в это время представителей кишечнополостных.
Палеозой. Палеозой (от греч. «древняя жизнь») - геологическая эра (570-230 млн. лет) со следующими периодами:
· кембрий (570-500 млн.лет)
· ордовик (500-440 млн. лет)
· силур (440-410 млн. лет)
· девон (410-350 млн. лет)
· карбон (350-285 млн. лет)
· пермь (285-230 млн. лет)
Для развития жизни в раннем палеозое (кембрий, ордовик, силур) характерно интенсивное развитие наземных растений и выход на сушу животных. Наступивший в конце силура горообразовательный период изменил климат и условия существования организмов. В результате поднятия суши и сокращения морей климат девона был более континентальный, чем в силуре. В девоне появились пустынные и полупустынные области; на суше появляются первые леса из гигантских папоротников, хвощей и плаунов. Новые группы животных начинают завоевывать сушу, но их отрыв от водной среды не был еще окончательным. К концу карбона относится появление первых пресмыкающихся - полностью наземных представителей позвоночных. Они достигли значительного разнообразия в перми из-за засушливого климата и похолодания. Так в палеозое произошло завоевание суши многоклеточными растениями и животными.
Мезозой. Мезозой (с греч. «средняя жизнь») - это геологическая эра (230-67 млн.лет) со следующими периодами:
· триас (230-195 млн.лет)
· юра (195-137 млн.лет)
· мел (137-67 млн.лет)
Мезозой справедливо называют эрой пресмыкающихся. Их расцвет, широчайшая дивергенция и вымирание происходят именно в эту эру. В мезозое усиливается засушливость климата. Вымирает множество сухопутных организмов, у которых отдельные этапы жизни связаны с водой: большинство земноводных, папоротники, хвощи и плауны. Вместо них начинают преобладать наземные формы, в жизненном цикле которых нет стадий, связанных с водой. В триасе среди растений сильного развития достигают голосеменные, среди животных - пресмыкающиеся. В триасе появляются растительноядные и хищные динозавры. Весьма разнообразны в эту эру морские пресмыкающиеся. Помимо ихтиозавров, в морях юры появляются плезиозавры. В юре пресмыкающиеся начали осваивать и воздушную среду. Летающие ящеры просуществовали до конца мела. В юре от пресмыкающихся возникли и птицы. На суше в юре встречаются гигантские растительноядные динозавры. Во второй половине мела возникли сумчатые и плацентарные млекопитающие. Приобретение живорождения, теплокровности были теми ароморфозами, которые обеспечили прогресс млекопитающих.
Кайнозой. Геологическая эра, в которую мы живем, называется кайнозой.
Кайнозой (от греч. «новая жизнь») - это эра (67 млн. лет - наше время) расцвета цветковых растений, насекомых, птиц и млекопитающих.
Кайнозой делится на два неравных периода: третичный (67-3 млн.лет) и четвертичный (3 млн.лет - наше время). В первой половине третичного периода широко распространены леса тропического и субтропического типа. В течение третичного периода от насекомоядных млекопитающих обособляется отряд приматов. К середине этого периода широкое распространение получают и общие предковые формы человекообразных обезьян и людей. К концу третичного периода встречаются представители всех современных семейств животных и растений и подавляющее большинство родов.
В это время начинается великий процесс остепнения суши, который привел к вымиранию одних древесных и лесных форм и к выходу других на открытое пространство. В результате сокращения лесных площадей одни из форм антропоидных обезьян отступали вглубь лесов, другие спустились с деревьев на землю и стали завоевывать открытые пространства. Потомками последних являются люди, возникшие в конце третичного периода.
В течение четвертичного периода вымирают мамонты, саблезубые тигры, гигантские ленивцы, большерогие торфяные олени и другие животные. Большую роль в вымирании крупных млекопитающих сыграли древние охотники.
Около 10 тысяч лет назад в умеренно теплых областях Земли наступила «неолитическая революция», связанная с переходом человека от собирательства и охоты к земледелию и скотоводству. Это определило видовой состав органического мира, который существует в настоящее время.