

Билет 8
Общие закономерности организации биосферы на Земле.
Биосфера — оболочка Земли, которая включает совокупность живых существ и в которой возможна постоянная жизнь.
Определение Биосферы, как особой оболочки Земли и само её название были предложены Зюссом (1873) в его работе «Лик Земли».
Вернадский создал учение о биосфере, показав, что живые организмы являются мощной биогеохимической силой, влияющей на геологические процессы (1926г.). Он считал, что биосфера состоит из:
1)живое вещество - растения, животные
2)биогенное вещество - каменный уголь, торф
3)косное вещество - горные породы неорганического происхождения и вода
4)биокосное вещество-взаимодействие живого и неживого (ил, почва, осадочные породы)
Две из многочисленных характеристик биосферы являются основополагающими: Всюду и всегда есть вода в жидком состоянии; В биосферу постоянно проникает солнечная радиация, представляющая единственный источник энергии.
Биосфера объединяет все экосистемы Земли и представляет собой глобальную экологическую систему - экосферу. Биогеоценозы являются элементарными структурами биосферы. Биосмасса и абиотические компоненты связаны сложными биогеохимическими процессами перераспределения энергии и вещества.
Границами биосферы являются пределы распространения живого на планете. Верхняя граница проходит примерно на высоте 20 км и ограничена слоем озона, в гидросфере организмы проникают на всю глубину Мирового Океана до 10-11км. В литосфере - 3,5-7,5 км.
В пределах биосферы В.И. Вернадский выделил «слой концентрации жизни» у поверхности Земли. Мощность этого слоя измеряется метрами, и только во влажных тропических лесах она достигает десятков метров.
Биосферу, как и любую другую систему, формируют не только внешние факторы, но и внутренние закономерности.
Согласно закону биогенной миграции атомов В.И. Вернадского, в общих химических процессах на поверхности планеты самое непосредственное участие принимает деятельность живого вещества.
Для живого вещества планеты неизбежна количественная стабильность, которая отражается в законе константности живого вещества Вернадского: количество живого вещества биосферы для данного геологического периода есть константа. Помимо количественной константности в живой природе наблюдается постоянное сохранение информационной и соматической (внутренней) структуры, хотя она медленно изменяется в процессе эволюции. Закон сохранения структуры биосферы – информационной и соматической – Ю. Голдсмит назвал первым законом экодинамики.
Э.Я. Колчинский выделяет следующие тенденции эволюции биосферы: постепенное увеличение ее общей биомассы и продуктивности; прогрессивное накопление аккумулированной солнечной энергии в поверхностных оболочках Земли; увеличение информационной емкости биосферы; усиление биогеохимических функций живого вещества и появление новых функций; расширение сферы действия биотического круговорота и усложнение его структуры.
Наличие ярко выраженного круговорота веществ, согласно закону глобального замыкания биогеохимического круговорота, – обязательное свойство биосферы любого этапа развития. В биосфере периодически происходит каскадная по иерархии систем глобальная перестройка. Одни виды за относительно короткое время безвозвратно исчезают, замещаются другими. Это периодическое убыстрение было отражено в виде принципа (правила) катастрофического толчка: природная катастрофа приводит к существенным эволюционным перестройкам, которые прогрессивны для биосферы, так как адаптируют ее к новым условиям среды. Поскольку существуют ускорения и замедления эволюции, действует и принцип прерывности и непрерывности развития биосферы: процесс медленного эволюционного изменения организмов закономерно прерывается фазами бурного развития и вымирания без переходных (палеонтологических) форм.
По утверждению В. Гранта в течение кайнозойского периода число видов оставалось примерно одинаковым, что отражается в законе константности числа видов в ходе стационарной эволюции биосферы: число народившихся видов в среднем равно числу вымерших, и общее видовое разнообразие в биосфере есть константа. Отсюда следует правило обязательности заполнения экологических ниш.
Общие закономерности организации биосферы
Биосфера – своеобразная оболочка Земли, содержащая совокупность живых организмов и ту часть веществ планеты, которая находится в непрерывном обмене с этими организмам. По В.И.Вернадскому, человеческий фактор становится главенствующим в процессе изменения биосферы. «Лик планеты – биосфера – химически резко меняется человеком сознательно и главным образом бессознательно. Меняется человеком физически и химически воздушная оболочка суши, все её природные воды», - писал он в своём научном труде «Химическое строение биосферы Земли и её окружения».
Очевидно, что все возрастающие социально-экономические потребности развития общества пришли в явное противоречие с ограниченными ресурсовоспроизводящими и жизнеобеспечивающими возможностями биосферы. Происходит истощение естественных ресурсов суши и океана, безвозвратная потеря различных видов растений и животных, загрязнение природной среды, деградация экосистем.
Возникла необходимость предотвращения разрушения биосферы, разработки мероприятий и осуществления контроля за их эффективностью, обеспечивающих устойчивость окружающей среды при возникающих антропогенных возмущениях.
Хозяйственная деятельность человека в процессе материального производства зачастую ведёт к разрушению природных систем – эстетических, исторических и информационных ценностей. В результате экологические требования по сохранению компонентов окружающей среды вступают в диалектические противоречия с хозяйственной деятельностью человека, направленной на удовлетворение его материальных потребностей.
Такое противоречие обостряется в случае уничтожения невосполнимых запасов, ценность которых возрастает по мере их исчерпания.
Сочетание хозяйственной деятельности с воспроизводством природных ресурсов и сохранением благоприятной среды жизни на Земле – необходимое условие природопользования.
Под воспроизводством природных ресурсов в природопользовании понимается рост природных богатств, а не простое их поддержание в неизменных масштабах. Системная совокупность всех природных ресурсов (материальных, энергетических и информационных) как факторов жизни общества в сочетании с человеческими ресурсами составляет интегральные ресурсы. Интегральный ресурс позволяет выявить зависимость качественных и количественных изменений одного из ресурсов от существующих перемен в качестве и количестве других ресурсов. Воспроизводство природных ресурсов принципиально невозможно без всемерной интенсификации и более глубокого использования природных благ (например, при замене одного вида энергии и качестве альтернативы другим).
При этом необходимо отличать воспроизводство природных ресурсов от их возобновления и восстановления, их экономического возмещения и замены, что практикуется в хозяйственной деятельности человека.
Предприятия-природопользователи выплачивают натуральную компенсацию государству как собственнику природных ресурсов за ущерб в природопользовании (например, восстановление леса, залужение склонов, рекультивация земель и т.д.), причём компенсация должна проводиться непосредственно в месте нанесения ущерба виновником.
При решении проблем обеспечения материальных потребностей человека следует учитывать локальные, региональные и глобальные в настоящем и обозримом будущем, наметив при этом приоритетные направления для их удовлетворения.
Приоритет в природопользовании должен отдаваться сохранности общемировых и национальных невосполнимых ценностей вопреки кажущейся сиюминутной экономической необходимости их утрат. Иначе социальные издержки могут оказаться неоправданно высокими и привести впоследствии к большим экономическим потерям.
С ростом хозяйственной деятельности человека происходит и возрастание антропогенной нагрузки на природную среду. Необходимо установить предел у природной среды в противостоянии антропогенным нагрузкам, и это поможет выявить эколого-экономический потенциал, характеризующий максимально допустимую антропогенную нагрузку на всю самоорганизованную совокупность природных систем. Нагрузка не должна приводить к необратимому разрушению совокупности систем, а также к сбоям в системных законах и резкому ухудшению её динамических качеств.
В качестве примера сбоев эколого-экономического потенциала можно привести некоторые прибрежные акватории морей, в которых в результате повышенных сбросов загрязнённых вод, нарушения естественных водотоков происходит замедление процессов естественного самоочищения.
Для восстановления частоты водоёмов потребуются более длительные сроки, а также денежные затраты для осуществления дополнительных мер по восстановлению жизненного цикла водоёмов. Эколого-экономический потенциал характеризует сохраняющуюся надёжность всех природных систем на всех уровнях.
Эколого-экономический потенциал должен учитываться вместе с природно-ресурсным потенциалом, который характеризует способность природных систем без ущерба для себя отдавать необходимую человеку продукцию или производить полезную работу. Он должен учитываться при планировании акций природопользования как экологическое ограничение в развитии хозяйства.
Экологическое ограничение – это ландшафтно-биологический или физико-химический «запрет», связанный с фактически предполагаемыми неблагоприятными воздействиями техногенных компонентов на среду жизни.
Экологическое ограничение для энергетики оценивается значением, близким по величине к энергетике глобального фотосинтеза (не более 20% от поступающей к поверхности Земли солнечной энергии). Преодолеть это экологическое ограничение практически невозможно, поскольку этой величины ограничивается предельная мощность развития энергетики на Земле.
Общая характеристика механизмов наследственности и изменчивости. Мутационная и модификационная изменчивость.
Модификационная (фенотипическая) изменчивость — изменения в организме, связанные с изменением фенотипа вследствие влияния окружающей среды и носящие, в большинстве случаев, адаптивный характер. Генотип при этом не изменяется. В целом современное понятие «адаптивные модификации» соответствует понятию «определенной изменчивости», которое ввел в науку Чарльз Дарвин.
Условная классификация модификационной изменчивости
По изменяющимся признакам организма:
морфологические изменения
физиологические и биохимические адаптации — гомеостаз (повышение уровня эритроцитов в горах и т. д.)
По размаху нормы реакции
узкая (более характерна для качественных признаков)
широкая (более характерна для количественных признаков)
По значению:
модификации (полезные для организма — проявляются как приспособительная реакция на условия окружающей среды)
морфозы (ненаследственные изменения фенотипа под влиянием экстремальных факторов окружающей среды или модификации, возникающие как выражение вновь возникших мутаций, не имеющие приспособительного характера)
фенокопии (различные ненаследственные изменения, копирующие проявление различных мутаций)— разновидность морфозов
По длительности:
есть лишь у особи или группы особей, которые подверглись влиянию окружающей среды (не наследуются)
длительные модификации — сохраняются на два-три поколения
Механизм модификационной изменчивости
Окружающая среда как причина модификаций
Модификационная изменчивость — это результат не изменений генотипа, а его реакции на условия окружающей среды. При модификационной изменчивости наследственный материал не изменяется, — изменяется проявление генов.
Под действием определенных условий окружающей среды на организм изменяется течение ферментативных реакций (активность ферментов) и может происходить синтез специализированных ферментов, некоторые из которых (MAP-киназа и др.) ответственны за регуляцию транскрипции генов, зависящую от изменений окружающей среды. Таким образом, факторы окружающей среды способны регулировать экспрессию генов, то есть интенсивность выработки ими специфических белков, функции которых отвечают специфическим факторам окружающей среды.
Например, за выработку меланина ответственны четыре гена, которые находятся в разных хромосомах. Наибольшее количество доминантных аллелей этих генов — 8 — содержится у людей негроидной расы. При воздействии специфической окружающей среды, например, интенсивного воздействия ультрафиолетовых лучей, происходит разрушение клеток эпидермиса, что приводит к выделению эндотелина-1 и эйкозаноидов. Они вызывают активацию фермента тирозиназы и его биосинтез. Тирозиназа, в свою очередь, катализирует окисление аминокислоты тирозина. Дальнейшее образование меланина проходит без участия ферментов, однако большее количество фермента обуславливает более интенсивную пигментацию.
Норма реакции
Предел проявления модификационной изменчивости организма при неизменном генотипе — норма реакции. Норма реакции обусловлена генотипом и различается у разных особей данного вида. Фактически норма реакции — спектр возможных уровней экспрессии генов, из которого выбирается уровень экспрессии, наиболее подходящий для данных условий окружающей среды. Норма реакции имеет пределы или границы для каждого биологического вида (нижний и верхний) — например, усиленное кормление приведет к увеличению массы животного, однако она будет находиться в пределах нормы реакции, характерной для данного вида или породы. Норма реакции генетически детерминирована и наследуется. Для разных признаков пределы нормы реакции сильно различаются. Например, широкие пределы нормы реакции имеют величина удоя, продуктивность злаков и многие другие количественные признаки), узкие пределы — интенсивность окраски большинства животных и многие другие качественные признаки.
Тем не менее, для некоторых количественных признаков характерна узкая норма реакции (жирность молока, число пальцев на ногах у морских свинок), а для некоторых качественных признаков — широкая (например, сезонные изменения окраски у многих видов животных северных широт). Кроме того, граница между количественными и качественными признаками иногда весьма условна.
Характеристика модификационной изменчивости
обратимость — изменения исчезают при смене специфических условий окружающей среды, спровоцировавших их
групповой характер
изменения в фенотипе не наследуются, наследуется норма реакции генотипа
статистическая закономерность вариационных рядов
затрагивает фенотип, при этом не затрагивая сам генотип.
Анализ и закономерности модификационной изменчивости
Вариационный ряд
Ранжированное отображение проявления модификационной изменчивости — вариационный ряд — ряд модификационной изменчивости свойства организма, который состоит из отдельных свойств видоизменений, размещенных в порядке увеличения или уменьшения количественного выражения свойства (размеры листка, изменение интенсивности окраски шерсти и т. д.). Единичный показатель соотношения двух факторов в вариационном ряде (например, длина шерсти и интенсивность ее пигментации) называется варианта. Например, пшеница, растущая на одном поле, может сильно отличаться количеством колосьев и колосков в силу различных показателей почвы, увлажненности на поле. Составив число колосков в одном колосе и количество колосьев, можно получить вариационный ряд в статистической форме:
Вариационный ряд модификационной изменчивости пшеницы
Число колосков в одном колосе 14 15 16 17 18 19 20
Количество колосьев пшеницы 2 7 22 32 24 8 5
Вариационная кривая
График среднеквадратичного отклонения, исходящий из вариационной кривой «модификационная изменчивость пшеницы»
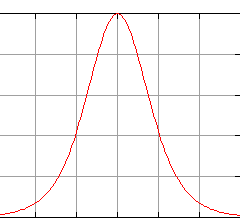
Графическое отображение проявления модификационной изменчивости — вариационная кривая — отображает как диапазон вариации свойства, так и частоту отдельных вариант. Из кривой видно, что наиболее распространены средние варианты проявления признака (закон Кетле). Причиной этого, по-видимому, является действие факторов окружающей среды на ход онтогенеза. Некоторые факторы подавляют экспрессию генов, другие же, наоборот, усиливают. Почти всегда эти факторы, одновременно действуя на онтогенез, нейтрализуют друг друга, то есть ни уменьшения, ни увеличения значения признака не наблюдается. Это и является причиной, по которой особи с крайними выражениями признака встречаются в значительно меньшем количестве, чем особи со средней величиной. Например, средний рост мужчины — 175 см — встречается в европейских популяциях наиболее часто.
При построении вариационной кривой можно рассчитать величину среднеквадратичного отклонения и, на основе этого, построить график среднеквадратичного отклонения от медианы — наиболее часто встречающуюся величину признака.
Модификационная изменчивость в теории эволюции
Дарвинизм
В 1859 году Чарльз Дарвин опубликовал свою работу на эволюционную тему под названием «Происхождение видов путём естественного отбора, или сохранение благоприятных рас в борьбе за жизнь». В ней Дарвин показал постепенное развитие организмов как результат естественного отбора. Естественный отбор состоит из такого механизма:
сначала появляется особь с новыми, совершенно случайными, свойствами (образованными вследствие мутаций)
потом она оказывается или не оказывается способной оставить потомство, в зависимости от этих свойств
наконец, если исход предыдущего этапа оказывается положительным, то она оставляет потомство и её потомки наследуют новоприобретённые свойства
Новые свойства особи формируются вследствие наследственной и модификационной изменчивости. И если наследственная изменчивость характеризуется изменением генотипа и эти изменения наследуются, то при модификационной изменчивости наследуется способность генотипа организмов изменять фенотип при воздействии окружающей среды. При постоянном воздействии одних и тех же условий окружающей среды на генотип могут отбираться мутации, чей эффект сходен с проявлением модификаций, и, таким образом, модификационная изменчивость переходит в наследственную изменчивость (генетическая ассимиляция модификаций). Примером может являться постоянный большой процент пигмента меланина в коже у негроидной и монголоидной расы по сравнению с европеоидной.
Дарвин назвал модификационную изменчивость определенной (групповой).
Определенная изменчивость проявляется у всех нормальных особей вида, подвергшихся определенному воздействию. Определенная изменчивость расширяет пределы существования и размножения организма.
Естественный отбор и модификационная изменчивость
Модификационная изменчивость тесно связана с естественным отбором. Естественный отбор имеет четыре направления, три из которых непосредственно нацелены на выживание организмов с разными формами ненаследственной изменчивости. Это стабилизирующий, движущий и дизруптивный отбор.
Стабилизирующий отбор характеризуется обезвреживанием мутаций и формирования резерва этих мутаций, что обуславливает развитие генотипа при постоянном фенотипе. Вследствие этого организмы со средней нормой реакции доминируют в неизменных условиях существования. Например, у генеративных растений сохраняется форма и размер цветка, которые отвечают форме и размеру насекомого, которое опыливает растение.
Дизруптивный отбор характеризуется раскрытием резервов с обезвреженными мутациями и последующим отбором этих мутаций для формирования новых генотипа и фенотипа, которые подходят под окружающую среду. Вследствие этого выживают организмы с крайней нормой реакции. Например, насекомые с большими крыльями имеют большую устойчивость к порывам ветра, тогда как насекомых того же вида со слабыми крыльями сдувает.
Движущий отбор характеризуется тем же механизмом, что и дизруптивный, однако он нацелен на формирование новой средней нормой реакции. Например, у насекомых появляется стойкость к химикатам.
Эпигенетическая теория эволюции
Согласно основным положениям эпигенетической теории эволюции, опубликованным в 1987 году, субстратом для эволюции является целостный фенотип — то есть, морфозы в развитии организма определяются воздействием условий окружающей среды на его онтогенез (эпигенетическая система). При этом формируется устойчивая траектория развития, основанная на морфозах (креод) — формируется устойчивая эпигенетическая система, адаптивная к морфозам. Эта система развития основана на генетической ассимиляции организмов (модификационном генокопировании), которая состоит в соответствии какой-либо модификации определенной мутации. То есть, это значит, что изменение активности конкретного гена может быть вызвано и изменением окружающей среды, и определенной мутацией. При действии новой окружающей среды на организм происходит отбор мутаций, которые приспосабливают организм к новым условиям, поэтому организм, вначале приспосабливаясь к среде с помощью модификаций, затем стает приспособленным к нему и генетически (двигательный отбор) — возникает новый генотип, на основе которого возникает новый фенотип. Например, при врожденном недоразвитии двигательного аппарата животных возникает перестройка опорных и двигательных органов таким образом, что недоразвитие окажется адаптативным. Далее этот признак закрепляется наследственно стабилизирующим отбором. Впоследствии возникает новый механизм поведения, направленный на приспособление к адаптации. Таким образом, в эпигенетической теории эволюции рассматривается постэмбриональный морфоз на основе особых условий окружающей среды как двигательный рычаг эволюции. Таким образом, естественный отбор в эпигенетической теории эволюции состоит из следующих стадий:
экстремальный фактор окружающей среды приводит морфозам, а морфозы — к дестабилизации онтогенеза
дестабилизация онтогенеза приводит к проявлению нестандартного (альтернативного, аномального) фенотипа, который наиболее отвечает сложившимся морфозам
при удачном соответствии альтернативного фенотипа происходит закрепленное генокопирование модификаций, которое ведет ко стабилизации онтогенеза и обуславливает направление естественного отбора — устанавливается новая норма реакции
далее, в ходе закрепления новых свойств путем генокопирования модификаций, формируются новые альтернативные пути развития, которые проявляются при следующей дестабилизации онтогенеза[1]
Таким образом, синтетическая и эпигенетическая теории эволюции достаточно различаются. Однако могут встречаться случаи, которые являются синтезом данных теорий — например, появления морфозов вследствие накопления нейтральных мутаций в резервах являются частью механизма как синтетической (мутации проявляются в фенотипе), так и эпигенетической (морфозы могут привести к генокопированию мдификаций, если изначальные мутации не детерминировали это) теорий.
Формы модификационной изменчивости
В большинстве случаев модификационная изменчивость способствует положительной адаптации организмов к условиям окружающей среды — улучшается реакция генотипа на окружающее и возникает перестройка фенотипа (например, увеличивается число эритроцитов у человека, поднявшегося в горы). Однако иногда, под влиянием неблагоприятных факторов окружающей среды, например, влиянием тератогенных факторов на беременных, возникают изменения фенотипа, похожие на мутации (не наследственные изменения, похожие на наследственные) — фенокопии. Также, под влиянием экстремальных факторов окружающей среды, у организмов могут появиться морфозы (например, расстройство двигательного аппарата вследствие травмы). Морфозы имеют необратимый и неадаптивный характер, а в лабильном характере проявления схожи со спонтанными мутациями. Морфозы принимаются эпигенетической теорией эволюции как основной фактор эволюции.
Длительная модификационная изменчивость
В большинство случаев модификационная изменчивость носит ненаследственный характер и является лишь реакцией генотипа данной особи на условия среды с последующим изменением фенотипа. Однако известны и длительные модификации, описанные у некоторых бактерий, простейших и многоклеточных эукариот. Для понимания возможного механизма длительной модификационной изменчивости рассмотрим сначала понятие генетического триггера.
Например, в оперонах бактерий содержатся, кроме структурных генов, два участка — промотор и оператор. Оператор некоторых оперонов находится между промотором и структурными генами (у других он входит в состав промотора). Если оператор связан с белком, который называется репрессором, то вместе они не дают двигаться РНК-полимеразе по цепи ДНК. У бактерий E. сoli можно наблюдать подобный механизм. При недостатке лактозы и избытке глюкозы вырабатывается белок-репрессор (Lacl), который присоединяется к оператору, не давая РНК-полимеразе синтезировать мРНК для трансляции фермента, который расщепляет лактозу. Однако при попадании лактозы в цитоплазму бактерии лактоза (вещество-индуктор) присоединяется к белку-репрессору, изменяя его конформацию, что приводит к диссоциации репрессора от оператора. Это обуславливает начало синтеза фермента для расщепления лактозы.
У бактерий при делении вещество-индуктор (в случае с E. coli — лактоза) передается в цитоплазму дочерней клетки и запускает диссоциацию белка-репрессора от оператора, что влечет за собой проявление активности фермента (лактазы) для расщепления лактозы у палочек даже при отсутствии этого дисахарида в среде.
Если оперона два и если они взаимосвязаны (структурный ген первого оперона кодирует белок-репрессор для второго оперона и наоборот), они образуют систему, которая называется триггером [1]. При активном состоянии первого оперона отключен второй. Однако под действием окружающей среды может быть заблокирован синтез белка-репрессора первым опероном, и тогда происходит переключение триггера: активным становится второй оперон. Такое состояние триггера может наследоваться следующими поколениями бактерий. Молекулярные триггеры могут обеспечивать длительные модификации и у эукариот. Это может происходить, например, путем цитоплазматического наследования включений цитоплазмы у бактерий при их размножении.
Эффект переключения триггеров можно наблюдать у неклеточных форм жизни, например, у бактериофагов. При внедрении в клетку бактерии при недостатке питательных веществ они остаются неактивными, внедряясь в генетический материал. При появлении благоприятных условий в клетке фаги размножаются и вырываются из бактерии — происходит переключение триггера вследствие изменения окружающей среды.
Мутационная изменчивость
Эволюционные преобразования организмов основываются на изменениях аппарата наследственности, приводящих к соответствующим перестройкам нормы реакции генотипа; такие изменения получили название мутаций. Мутации могут возникнуть (и неизбежно возникают в определенном проценте случаев) в процессе редупликации генома, предшествующем клеточному делению, или в процессе самого клеточного деления. Точные причины естественно происходящих (спонтанных) мутаций обычно неизвестны, но, очевидно, они кроются в каких-то нарушениях нормального режима сложных процессов редупликации и расхождения хромосом, связанных в конечном итоге с внешними воздействиями на клетку. Возникновение спонтанных мутации связывают, например, с мутагенной активностью некоторых метаболитов (продуктов обмена веществ) или с термическими флюктуаииями атомов в процессах редупликации и т. п.
Размножение и связанная с ним редупликация наследственного аппарата представляют собой важнейшие и неотъемлемые свойства живых организмов. Поскольку эти сложные процессы протекают на фоне постоянных изменений внешней среды и поскольку всякий механизм и всякая структура обладают конечной степенью устойчивости, нарушения процесса редупликации, ведущие к мутациям, совершенно неизбежны. Мутации представляют собой элементарные изменения, являющиеся материалом для действия естественного отбора. Неизбежный результат естественного отбора - эволюционные изменения организмов. Таким образом, как отметил Н.В.Тимофеев-Ресовский, неизбежность эволюции следует из основных свойств живой материи.
Хотя стимулом мутаций являются изменения внешних условий, между этими последними и происходящими под их влиянием мутациями нет адекватности. Это означает, что возникающие мутации сами по себе не являются приспособлениями к изменениям условий, вызвавшим их появление. Аппарат наследственности не обладает и принципиально не может обладать свойством целесообразно перестраиваться в ответ на изменения внешней среды. Это отчетливо проявляется при искусственном мутагенезе, когда внешние воздействия, вызвавшие возникновение мутаций, точно известны. Например, хорошо известно мутагенное действие проникающей радиации, но возникшие под ее влиянием мутации могут затрагивать самые различные особенности организмов (цвет глаз, степень развития и особенности строения крыльев у мух и т. п.), которые в подавляющем большинстве случаев никак не связаны с защитой организма от радиации. Точно так же действуют и другие, более "мягкие" мутагены (различные химические факторы, изменения температуры и др.), причем разные мутагены могут вызывать у данного вида организмов одни и те же мутации.
Этот факт важен в двух отношениях. С одной стороны, он подчеркивает неопределенный, неприспособительный характер мутаций, с другой - многократное появление у разных особей данного вида организмов одних и тех же мутаций. Из этого следует чрезвычайно важный вывод: всякий генотип имеет хотя и большие, но вполне конкретные возможности мутационных изменений, определенный спектр изменчивости. Одни и те же мутации с определенной регулярностью появляются в пределах популяций данного вида вновь и вновь. И с другой стороны, некоторые изменения, кажущиеся столь же вероятными, никогда у этого вида не наблюдаются. Например, среди различных видов мух-Дрозофил, вероятно наиболее изученных в генетическом отношении животных, никогда не наблюдались особи с зелеными или синими глазами. Поскольку генетиками изучено уже около двух миллиардов (!) мух, вероятность обнаружения указанных мутаций в будущем выглядит ничтожной. По-видимому, в геноме дрозофил нет каких-то предпосылок, необходимых для возникновения таких мутаций, хотя подобные окраски глаз нередко наблюдаются у других видов насекомых. Для дрозофил эти мутации являются по каким-то причинам невозможными ("запрещенными").
Мутации могут происходить на разных уровнях организации наследственного аппарата: на уровне полного хромосомного набора (генома) клетки, на уровне его морфологически обособленных частей -- хромосом и на уровне его структурно-функциональных единиц - генов.
Геномные мутации представляют собой изменения количества хромосом, которые могут происходить либо путем кратного увеличения числа целых (гаплоидных) наборов хромосом, либо посредством изменения числа отдельных хромосом в геноме, в результате чего общее число хромосом становится не кратным гаплоидному.
Увеличение числа целых хромосомных наборов (плоидности) в клетке называется полиплоидизацией, изменение числа отдельных хромосом в геноме - анеуплоидией, или гетероплоидией. Фенотипический эффект геномных мутаций основан на взаимодействии генов или их комплексов, расположенных в соответственных (гомологичных) хромосомах, число которых изменилось в результате данной мутации. Вероятно, это взаимодействие осуществляется через посредство продуктов белкового синтеза (ферментов) или РНК, транскрибированной с ДНК соответствующих хромосом.
Геномные мутации обычно приводят к стерильности мутантов в результате невозможности осуществления полового процесса с представителями родительской формы, сохранившими нормальное количество хромосом. Поэтому у животных относительно редко геномные мутации дают начало новым формам - это возможно только при способности данного вида размножаться бесполым способом, или посредством партеногенеза, или гиногенеза (гиногенез - способ размножения, при котором сперматозоид проникает в яйцеклетку и стимулирует ее развитие, но его ядро не сливается с ядром яйца и не участвует в развитии зародыша). Среди растений бесполое размножение распространено гораздо шире, чем среди животных. В связи с этим у растений полиплоидизация, вероятно, играла существенную роль в эволюционном процессе.
Обычно различают автополиплоидию и аллополиплоидию. Автополиплоиды образуются посредством увеличения числа хромосомных наборов у отдельных особей одного вида. У автополиплоидов каждая хромосома представлена одним и тем же числом гомологов, например в клетках тетраплоидов имеется по четыре гомологичные хромосомы, у октоплоидов - по восемь и т. д. Ал-лополиплоиды образуются посредством гибридизации организмов разных видов. У аллополиплоидов соответствующие хромосомы уже в определенной степени утратили свою гомологичность. Поэтому процесс мейоза, в котором осуществляется редукция числа хромосомных наборов перед половым процессом, у аллополиплоидов обычно нарушен. Аллополиплоиды могут сохраниться как новый вид, если способны к бесполому размножению.
Половой процесс может восстановиться у полиплоидной формы, когда в популяции накапливается достаточное количество полиплоидных особей. У аллополиплоидов, кроме того, необходимо еще дополнительное кратное удвоение числа хромосом (посредством эндомитоза в соматических клетках или нерасхождения хромосом при образовании гамет). (Эндомитоз - удвоение числа хромосом внутри ядра без образования веретена деления клетки). Тогда в аллополиплоидном наборе появляются гомологичные хромосомы, и в дальнейшем мейоз может протекать нормально. Известно несколько примеров возникновения новых форм растений этим путем; среди них можно упомянуть рафанобрассику - межродовой гибрид, полученный Г.Д.Карпеченко путем скрещивания редьки (Raphanus sativus) с капустой (Brassica oleracea).
Хромосомные мутации являются структурными перестройками отдельных хромосом, изменяющими расположение в них генов, фенотипическое проявление подобных мутаций зависит от изменений пространственной локализации генов в хромосомах (эффект положения). Выделяют несколько основных типов хромосомных мутаций: а) транслокация - взаимный обмен участками между двумя разными (не гомологичными друг другу) хромосомами; б) делеция - утрата хромосомой какого-либо ее участка; в) инверсия - поворот внутри хромосомы какого-либо ее участка на 180°; г) дупликация - удвоение тех или иных участков хромосомы; д) транспозиция - перестановка генов внутри хромосомы.
Среди указанных типов хромосомных мутаций особую роль играют дупликации, которые (так же как и полиплоидизация) позволяют увеличить количество генетического материала в клетках. Как подчеркнул С. Оно, возникновение избыточности генетического материала может играть важную роль в эволюционных перестройках генетической системы вида, открывая новые возможности для прогрессивной эволюции, в частности для развития качественных новообразований. Дупликации ведут к появлению в наследственном аппарате генов-дублеров, отвечающих за одну и ту же функцию. В таких дублерах могут в скрытой форме сохраняться и накапливаться новые генные мутации. С другой стороны, инверсии способствуют образованию устойчивых генных комплексов (групп сцепления), которые редко разрушаются кроссинговером при мейозе и обеспечивают совместное наследование различных признаков.
Наконец, генные или точковые мутации представляют собой изменения самих генов, т.е. элементарных функциональных единиц аппарата наследственности. Сущность генных мутаций заключается в изменении порядка пуриновых и пиримидиновых оснований в молекуле ДНК. Для возникновения одной точковой мутации достаточно изменения хотя бы одной пары оснований. Если такие мутации происходят в структурных генах, может измениться порядок считывания наследственной информации при транскрипции ("мутация рамки" - смешение рамки считывания наследственной информации), что приведет к изменению порядка аминокислот в полипептидной цепи, т. е. к синтезу нового типа белковых молекул. Мутации генов-регуляторов изменяют соотношения между репрессированными и активными структурными генами, что неизбежно влечет за собой изменения белковых комплексов, синтезируемых в различных типах соматических клеток, несмотря на оставшиеся неизменными структурные гены.
Генные мутации являются основным источником наследственной изменчивости организмов. В результате мутирования возникают различные варианты генов - аллели, обусловливающие разные состояния контролируемого данным геном признака (или признаков). Число аллельных состояний отдельных генов может быть очень большим (более ста).
Поскольку мутационная изменчивость неопределенна по отношению к характеру изменений внешней среды, вызвавших появление мутаций, подавляющее большинство возникших наследственных изменений не улучшают, а ухудшают приспособленность мутантных организмов, и тем в большей мере, чем больше масштаб фенотипического проявления мутации. Поэтому в среднем наибольшие шансы на сохранение имеют мутации с наименьшим фенотипическим выражением ("малые мутации"), которые и служат основным материалом эволюционных изменений.