

-
Окислительные реакции, катализируемые десатуразами. Строение и локализация, роль.
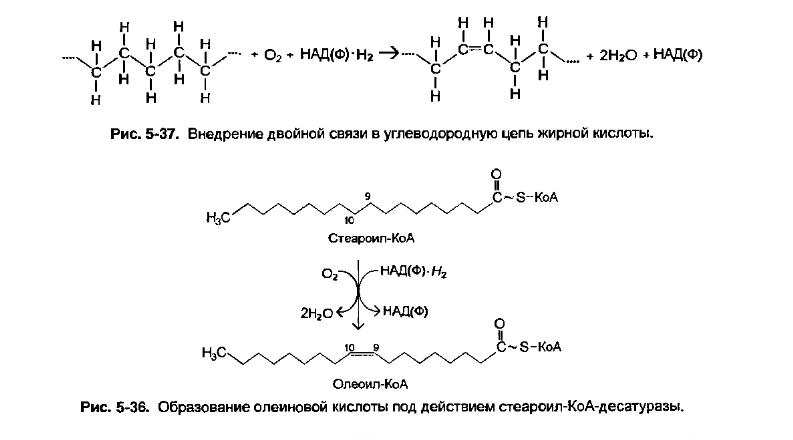
Название свое ферменты этой группы получили по способности превращать насыщенные жирные кислоты в ненасыщенные (и далее в полиненасыщенные). Образование двойной связи они обеспечивают путем отнятия двух атомов водорода у соседних углеродных атомов. При этом отнимаемый водород передается прямо на кислород. Но, в отлнчие от оксидаз- ных реакций, здесь происходит не двухэлектронное восстановление молекулы кислорода, а четырехэлектронное. Поэтому образуется не перекись водорода, а две молекулы воды. Недостающую пару электронов (и протонов) де- сатуразы получают от НАД(Ф)-//г- (с учетом того, что формируемая двойная связь имеет, как правило, ^с-конфигурацию).
Полнота восстановления 02 обеспечивается тем, что десатураза (несущая 1 атом неге- мового железа) - это не одиночный белок, а составная часть комплексов, встроенных в мембрану ЭР. В такой комплекс входят также цитохром Ь5 и цитохром(Ь$)-редуктаза, которая содержит ФАД и использует НАД(Ф)Я2 для восстановления железа цитохрома Ъ5_
Важная роль десатураз обусловлена тем, что биосинтез любой жирной кислоты протекает путем поэтапной конденсации двухуглеродных фрагментов (происходящих из ацетил- КоА), приводя к образованию полностью насыщенной углеводородной цепи (с группой -СООН на одном из концов). И только затем возможно внедрение двойных связей, что и осуществляют десатуразы. Жирные кислоты вступают в реакцию только в активированном состоянии, т.е., в форме соединения с коферментом А (ацил-КоА).
В отличие от растений клетки животного организма имеют очень ограниченную способность формировать двойную связь в молекулах жирных кислот. В отношении насыщенных известен только один фермент - стеарош- КоА -десатураза. открытая в печени как интегральный белок мембраны ЭР. Она относится к Д9-десатуразам, ибо внедряет двойную связь в положении 9 (т.е., между 9-м и 10-м атомами углерода, считая от карбоксильной группы). При этом стеариновая кислота С(18):0 превращается в олеиновую С(18):1 (рис. 5-38). Аналогичным образом стеароил-КоА-десатураза действует и на ряд других насыщенных жирных кислот, преобразуя их в моноеновые. В частности, она превращает пальмитиновую кислоту С(16):0 в пальмитолеиновую С(16):1
Полиеновые кислоты должны поступать в организм животных извне. Точнее, из всех жирных кислот незаменимыми компонентами пищн являются линолевая и а-линоленовая (содержащие по 18 углеродных атомов). Как отмечено в табл. 3.1, первая содержит две двойных связи (в позициях 9 и 12), а вторая - три (в положениях 9, 12 и 15). Обе они служат материалом для создания еще более ненасыщенных жирных кислот, которые необходимы для построения липидов биологических мембран и, кроме того, для образования сигнальных молекул из группы эйкозаноидов.
сначала линолевая и а-линоленовая кислоты подвергаются десатурации в положении 6, что превращает их соответственно в триеновую у-линоленовую кислоту и в ее тетраеновый аналог. В обоих случаях процесс катализирует линолеош-КоА-десатураза. По строению, локализации и механизму действия она очень близка стеароил-КоА-десатуразе, но специализирована на внедрении двойной связи в позиции 6 (т.е., является Дб~десатуразой). Образовавшиеся продукты могут подвергаться удлинению специальной системой ЭР, обозначаемой как элонгаза. Как и при биосинтезе насыщенных жирных кнслот, это происходит путем наращивания карбоксильного конца молекулы двухуглеродным фрагментом, происходящим из ацетил-КоА. Такое удлинение отодвигает от группы -СООН (на два номера) положение тех двойных связей, которые уже имелись в молекуле. В итоге С 18-кислоты преобразуются в 20-углеродные (эйкозано- вые), а двойные связи смещаются из положений 6, 9, 12... в позиции 8, 11, 14—, соответственно. Последующая десатурация по 5-му углеродному атому приводит к превращению исходной линолевой кислоты в 5,8,11,14-эйко- затетраеновую (арахидоновую кислоту), а а-линоленовой - в 5,8,11,14,17-эйкозапента- еновую. Пути превращения последних в биологически активные молекулы будут рассмотрены в разделах
-
Витамин В3 PP
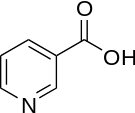
Никотиновая кислота
B5
Ниацин
Участвует в процессах клеточного дыхания (перенос H+ и e–). Регулирует секреторные и мо-торные функции желудочно-кишечного тракта и печени. Вхо-дит в состав НАД и НАДФ.
Мясо, печень, почки, рыба, дрожжи.
Признаки недост: Воспаление кожи (пеллагра), расстройства желудочно-кишечного тракта.
Билет №9