

-
Биоценоз экосистемы
-
Динамика биоценоза как результат межвидовых взаимодействий
Понятие о биоценозе, введенное К. Мёбиусом в 1877 г., сыграло важную роль в становлении системной концепции в экологии, сосредоточивая внимание исследователей на взаимодействиях между видами и возникающими вследствие этого целостными свойствами биологических сообществ. В отношении биоценоза сложились разные представления – от точки зрения Ф. Клементса (1928) и его последователей на биотическое сообщество как на организм высшего порядка (суперорганизм) до практического отрицания объективных оснований для концепции биоценоза в работах Л. Г. Раменского (1924), Г. Глизона (1926) и других исследователей.
В нашем понимании биоценоз – это совокупность всех популяций биологических видов, принимающих существенное участие в функционировании данной экосистемы. То есть биоценоз – это живая часть экосистемы.
В каждый момент времени t биоценоз имеет определенное видовое богатство (видовую насыщенность), которое измеряется числом входящих в него популяций различных видов – n(t), и количественный состав, характеризуемый набором из n(t) чисел x1(t),…,xn(t)(t), обозначающих численности или плотности этих популяций.
Среди разнообразных группировок видов в пределах биоценоза наибольшее теоретическое и практическое значение имеют консорции, выделяемые на основе трофических и топических отношений, и гильдии, отражающие функциональную близость между видами.
Консорцией называется совокупность видов, связанных пищевыми или прочими (топическими, фабрическими и др.) связями с некоторым видом, называемым эдификатором или детерминантом консорции, в качестве которого обычно выступает растение-автотроф. Гильдия определяется как группа видов, использующих один и тот же класс ресурсов среды одинаковым способом. Таким образом, понятие «гильдия» и «трофическая группировка» оказываются синонимами.
-
Видовое разнообразие стационарных биоценозов
С каждым из n видов биоценоза сопоставим некоторый показатель значимости pi (i = 1,2,…,n), рассматриваемый как мера относительного доминирования или участия данного вида в сообществе или в его функционировании. В качестве показателя значимости вида может использоваться процентная доля вида в первичной продукции сообщества или доля дыхания данного вида в общем дыхании видов биоценоза. При рассмотрении сообществ, составленных из особей сравнимого размера, биомассы и метаболизма, в качестве показателя значимости (доминирования) допустимо использовать просто индекс относительного обилия, иногда выражаемый в процентах.
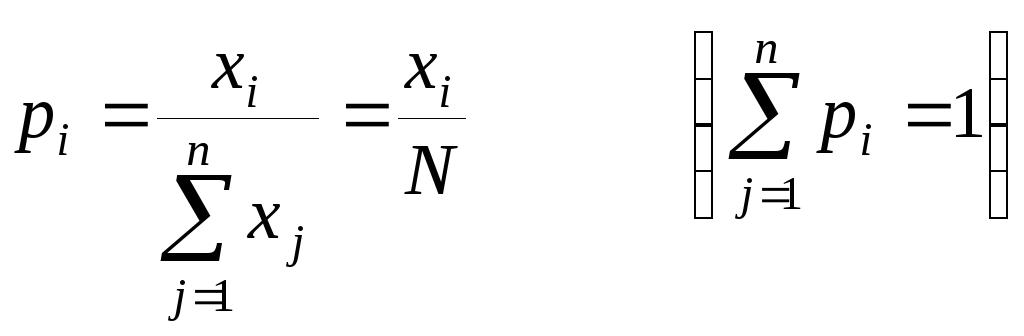 .
.
Из общего числа видов сообщества в целом обычно лишь немногие бывают доминирующими, то есть имеют значительную численность (большую биомассу, продуктивность и т.п.); большая же часть относится к редким видам. Правда, иногда доминантные виды отсутствуют, а имеется большое число видов со средним значением показателя значимости.
Видовое разнообразие слагается из двух компонентов:
1)
видового
богатства,
которое характеризуется общим числом
имеющихся видов
![]() где
где
![]() ;
;
2) выравненности, основанной на относительном обилии или другом показателе значимости вида и положении его в структуре доминирования.
Видовое разнообразие увеличивается при увеличении размеров площади и при продвижении от высоких широт к экватору. В сообществах же, подвергающихся стрессовым воздействиям, видовое разнообразие невелико; но, кроме того, оно может снижаться в результате конкуренции в старых сообществах, существующих в стабильной физической среде.
Большую роль играют два других типа разнообразия:
1) структурное разнообразие;
2) генетическое разнообразие, то есть поддержание генотипических гетерозиготности (присутствие в хромосоме доминантных и рецессивных признаков), полиморфизма и другой генотипической изменчивости, которая вызвана адаптационной необходимостью в природных популяциях.
Уменьшение видового и генотипического разнообразия, происходящее вследствие деятельности человека, ставит на грань риска возможность будущих адаптаций как в природных экосистемах, так и в агроэкосистемах.
Д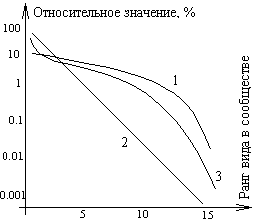 ля
изучения видового разнообразия принято
строить кривую
относительного доминирования-разнообразия,
введенную в рассмотрение Уиттэкером
(1965): по оси x
(абсцисс) отложена ранжированная
последовательность номеров видов i
от наиболее значимого до наименее
значимого, а по оси y
(ординат) – величина показателя
зависимости pi
соответствующего вида, обычно в
логарифмическом масштабе.
ля
изучения видового разнообразия принято
строить кривую
относительного доминирования-разнообразия,
введенную в рассмотрение Уиттэкером
(1965): по оси x
(абсцисс) отложена ранжированная
последовательность номеров видов i
от наиболее значимого до наименее
значимого, а по оси y
(ординат) – величина показателя
зависимости pi
соответствующего вида, обычно в
логарифмическом масштабе.
Выделяют три основных типа поведения кривой относительного доминирования, соответствующих различным типам взаимодействий между видами в сообществе.
Кривая 1 отвечает ситуации, когда члены сообщества находятся в сильной зависимости от некоторого ресурса, распределенного вдоль одномерного континуума, причем имеет место случайное, но без пересечений распределение частных ниш видов вдоль координатной оси данного ресурса (так называемая модель «разломанного стержня» Р. Мак-Артура). Показатель значимости в этом случае ведет себя в соответствии с формулой
![]() .
.
Подобный тип рангового распределения наиболее часто встречается в немногочисленных по числу видов группировках таксономически близких животных в четко ограниченных однородных биоценозах, как, например, группировка птиц, гнездящихся на ограниченном участке леса, а также справедлива для высших животных с активной конкуренцией, устойчивой численностью и высокой продолжительностью жизни.
Кривая 2, представляющая собой прямую, изображает случай так называемого «геометрического» распределения и соответствует следующей схеме относительного доминирования: первый вид занимает половину доступного пространства ниш, второй – половину оставшегося пространства (25% исходного) и т.д. В более общем случае существует фиксированное число с (0 с 1), такое, что
p1
= cp,
где
![]()
p2 = c(p - p1) = c(1 - c)p , p3 = c(p - p1 - p2) = c(1 - c)2 p
……………………
pi = c(1 - c)i-1 p
или в логарифмическом виде
log pi = (i - 1) log (1 - c) + log c + log p ,
откуда видно, что величина pi как функция ранга i изображается на полулогарифмической шкале прямой линией с угловым коэффициентом log (1 - c). Геометрическое распределение относительного доминирования обнаруживается в группировках, состоящих из небольшого числа видов, которые состоят в жесткой конкуренции за ограниченные ресурсы, нередко в суровых условиях внешней среды. Получаемую круто падающую прямую 2 можно рассматривать в качестве альтернативы для относительно пологой кривой 1, соответствующей распределению Мак-Артура.
Эти две кривые представляют крайние случаи, обычно же распределение в природе имеет вид промежуточной S-образной кривой 3, что указывает на более сложный характер дифференциации и перекрывания ниш. Распределение, изображаемое кривой 3, называется логнормальным. Это связано с тем, что гистограмма распределения логарифмов ln pi по октавам (диапазонам значений pi, отличающихся на границах в 2 раза) приближается по форме к нормальному распределению. Значения nj (число точек, попавших в данную октаву j) для различных октав с номерами j = …,-2,-1,0,1,2,… (где 0 – октава с наибольшим числом точек) могут быть аппроксимированы выражением
![]() ,
,
г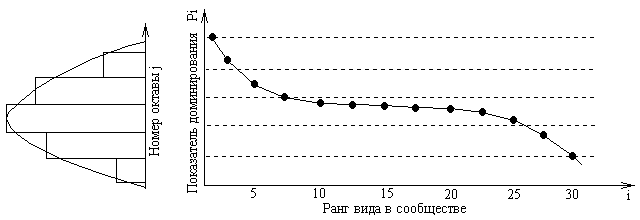 де
– мера рассеяния относительно модального
класса j=0,
которая для большинства анализируемых
видовых группировок находится в пределах
15.
де
– мера рассеяния относительно модального
класса j=0,
которая для большинства анализируемых
видовых группировок находится в пределах
15.
Логнормальное распределение относительного доминирования характерно для сообществ с высокой видовой насыщенностью, в условиях, когда «успех» того или иного вида определяется большим числом относительно независимых и однородных по силе влияния факторов. В этом случае ниши многомерны и перекрываются.
В общем расположение видов по степени значимости кривая доминирования не только точно отражает богатство и относительное обилие видов как компонентов видового разнообразия, но и объясняет, каким образом подразделяется пространство ниш. Чем выше кривая и чем более она уплощена, тем больше при данном числе видов общее разнообразие. Чем круче идет кривая, тем меньше общее разнообразие и сильнее доминирование одного или нескольких видов. В стрессовых ситуациях независимо от того, вызваны ли они естественными причинами (погодными условиями) или антропогенным воздействием, кривая становится более крутой. Таким образом, кривую доминирования можно использовать для оценки влияния нарушений на видовую структуру.
Кроме видового богатства и типа рангового распределения для характеристики сообществ используется так называемый показатель (индекс) разнообразия, который количественно отражает не только общее число видов, но и особенности количественного состава сообществ. К наиболее часто употребимым относят показатель разнообразия по Симпсону
![]()
показатель разнообразия по Шеннону
![]()
и показатель выравненности Пиелу
E = H / ln n .
Оба
показателя разнообразия обладают тем
свойством, что они принимают максимальное
значение при равенстве долей всех видов
в сообществе, то есть при p1
= p2
= … = pn
= 1/n
(![]() )
)
![]()
![]() .
.
С другой стороны, когда доля какого-нибудь одного вида, например i-го, стремится к 1, а всех остальных – к нулю, оба показателя также стремятся к нулю:
![]() ,
,
![]()
Из двух обобщенных индексов показатель Симпсона придает обычным видам больший вес, поскольку при возведении в квадрат малых значений pi получаются очень малые величины. Индекс Шеннона придает больший вес редким видам. Так как индекс Шеннона заимствован из теории информации и представляет собой формализацию, которая широко используется при оценке сложности и содержания информации в любых типах систем, он лучше всего подходит для целей сравнения в тех случаях, когда нас не интересуют компоненты разнообразия по отдельности.
При построении Ю. Одумом гистограммы для показателя разнообразия Симпсона для разных биоценозов выяснилось, что полученная гистограмма оказалась бимодальной: выделилась одна большая группа биоценозов с низким разнообразием (0.05 – 0.2) и другая большая группа со сравнительно высоким разнообразием (0.7 – 0.85). Во всей выборке не оказалось биоценоза с примерно равными значениями показателей относительного доминирования (p1 = … = pn = 1/n), когда индекс разнообразия приближается к своему максимальному значению, то есть к 1.
В группу с низким биотическим разнообразием попали: 1) экосистемы, находящиеся в состоянии стресса под действием внешних сил или притоков вещества или энергии, как в случае загрязненных рек или заливов, или экосистемы, активно регулируемые человеком, как посевы сельскохозяйственных культур или лесные плантации; 2) экосистемы, постоянно получающие большие количества высококачественной энергии или полезных веществ. Биоценозов со средним (около 0.5) разнообразием оказалось сравнительно мало.
Группы с высоким разнообразием образованы многими естественными экосистемами, не получающими извне концентрированной энергии или биогенных материалов и живущими только за счет солнца, например, степи, леса и большие озера.
Эти данные свидетельствуют о том, что видовое богатство и разнообразие биоценозов формируется в результате приспособления к силе и количественному составу приходящего энергетического потока. «Стратегия» природы состоит в увеличении разнообразия, но только до пределов, пока это не приводит к снижению эффективности использования ресурсов биоценоза. Следовательно, существует некоторый оптимум разнообразия, определяемый эффективностью экосистемы как энергетической машины. При больших внешних энергетических субсидиях в экосистеме эффективна простая, но специализированная структура с низким видовым разнообразием, способная использовать эти благоприятные условия более эффективно, чем менее специализированная совокупность видов.
В условиях ограниченного энергетического притока более выгодным для поддержания стационарного состояния экосистемы представляется высокий уровень разнообразия: порядка 0.7÷0.8. В этом случае достигается определенная надежность использования дефицитного источника энергии благодаря наличию многих альтернативных путей ее обесценивания.