


ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Возбудимыетканииихсвойства
Возбудимыми называются ткани, которые в ответ на раздражитель способны отвечать специфической формой активности: возникновением электрического потенциала или другими явлениями. К ним относятся нервная, мышечная и железистая ткани. Возбудимым тканям присущи следующие свойства:
–возбудимость – способность возбуждаться;
–проводимость – способность проводить нервный импульс (скорость проведения у нерва может достигать 120 м/с, т.е. 600 км/ч);
–сократимость – способность развивать силу или напряжение при возбуждении;
–лабильность – функциональная подвижность, т.е. способность к ритмической активности (нерв способен в 1 с генерировать до 1000 нервных импульсов);
–секреторная активность.
Сущность процесса возбуждения заключается в следующем. Все клетки организма ограничены от внешнего пространства двухслойной плазматической мембраной. Матрикс мембраны представлен главным образом фосфолипидами, гидрофильные группы которых обращены к водной среде, а гидрофобные углеводородные цепи располагаются в два ряда и образуют безводную липидную фазу, способную растворять неполярные молекулы (рис. 2). Разбросанные среди фосфолипидов в почти равном с ними количестве молекулы холестерола стабилизируют мембрану. Главными функциональными элементами, погруженными в билипидный слой, являются белки. Одни белки пронизывают мембрану, другие закреплены в каком-то одном слое. Обычно белки ориентированы так, что их гидрофобные группы погружены в липидную мембрану, а полярные гидрофильные группы находятся на поверхности мембраны, контактируя с межклеточной жидкостью. Многие белки наружной поверхности мембраны представлены гликопротеидами.
Цитозоль клетки и межклеточная жидкость электронейтральны.
Физиология. Конспект лекций |
-18- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Возбудимые ткани и их свойства
Рис. 2. Твердокаркасная модель плазматической мембраны
Однако все клетки организма имеют электрический заряд, обусловленный неодинаковой концентрацией катионов и анионов в наружном и внутреннем примембранном слоях клетки. Внутренний примембранный слой имеет отрицательный заряд, наружный – положительный. Наиболее распространенным катионом во внеклеточной среде является Na+, во внутриклеточной – K+ (рис. 3). При этом наибольший градиент внутри- и внеклеточной концентрации имеет место для ионов Ca2+, концентрация свободных ионов которого внутри клетки ниже в 25 000 раз, чем снаружи.
Рис. 3. Распределение ионов между вне- и внутриклеточной средами в двигательных нейронах спинного мозга (ммоль/л-1)
Физиология. Конспект лекций |
-19- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Возбудимые ткани и их свойства
Различная концентрация анионов и катионов является следствием неодинаковой проницаемости плазматической мембраны для разных ионов. Проницаемость плазматической мембраны определяется наличием в ее составе разных ионных каналов, размерами каналов и частиц, растворимостью частиц в мембране (клеточная мембрана проницаема для растворимых в ней липидов и непроницаема для пептидов). При действии раздражителя на клетку возбудимой ткани изменяется ионная проницаемость плазматической мембраны. В результате ионы быстро перемещаются через мембрану согласно электрохимическому градиенту (например, положительные ионы движутся в сторону избыточного отрицательного заряда и меньшей концентрации данного иона). Это и есть процесс возбуждения.
Транспортвеществчерезбиологическиемембраны
Главными функциональными элементами плазматической мембраны, обеспечивающими транспорт различных ионов, являются интегральные и периферические белки, погруженные в относительно инертный слой билипидов. Это белки, играющие роль ферментов, переносчиков, ионных каналов и рецепторов. Различают пассивный и активный транспорт, эндо- и экзоцитоз.
Пассивный транспорт представлен процессами простой и облегченной диффузии, обеспечивающей перенос веществ на небольшие расстояния. Простая диффузия происходит в соответствии с законом Фика – по химическому или электрохимическому градиенту, т.е. без затраты энергии (рис. 4). Такой тип диффузии характерен для воды и растворенных в ней газов, жирорастворимых веществ, а также некоторых полярных молекул небольшого размера. Облегченная диффузия происходит с участием белков-переносчиков или через специализированные ионные каналы.
Многие сахара, аминокислоты и нуклеотиды преодолевают мембрану с помощью транспортных белков, внутри которых имеется заполненный водой канал диаметром менее 1 нм. Связываясь с переносимой молекулой, белкипереносчики способствуют ее переносу по градиенту концентрации, т.е. без затраты энергии.
Физиология. Конспект лекций |
-20- |
ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Транспорт веществ через биологические мембраны
Рис. 4. Транспорт веществ через плазматическую мембрану
Рис. 5. Строение ионного канала
Ионные каналы представляют собой крупные интегральные белки, образующие центральную, наполненную водой пору, имеющую устье, селективный фильтр, ворота и механизм управления воротами (рис. 5). Проводимость каналов зависит от концентраций ионов около устья канала и проницаемости каналов. Большинство ионных каналов обладает селективностью, т.е. способностью пропускать только определенные ионы. По этому признаку различают натриевые, калиевые, кальциевые, хлорные каналы.
Селективность каналов обеспечивается размерами и геометрией поры (диаметром устья и селективного фильтра), размерами иона и его гидратной оболочки, зарядом иона и зарядом внутренней поверхности канала (например, внутри катионных каналов расположены анионы). Для того чтобы пройти через ионный канал, каждый ион должен освободиться от гидратной оболочки. При этом слишком большой ион не может войти в канал, а слишком маленький не способен отдать гидратную оболочку в селективном фильтре. Передвижение ионов в канале обеспечивается химической движущей силой,
Физиология. Конспект лекций |
-21- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Транспорт веществ через биологические мембраны
которая определяется разностью концентраций ионов внутри и снаружи клетки, и электрической движущей силой, зависящей от потенциала на мембране.
Активный транспорт веществ через биологические мембраны требует определенных энергетических затрат. Различают первично-активный транспорт и вторично-активный транспорт. Первично-активный транспорт широко представлен в организме. Это натрий-калиевый насос, кальциевый насос, натрий-водородный ионообменник, натрий-кальциевый ионообменник и т.д. Суть данного вида транспорта через биологические мембраны состоит в том, что в мембране имеется переносчик, обладающий АТФ-азной активностью, т.е. способный расщеплять АТФ на АДФ и фосфат и высвобождать энергию, необходимую для переноса вещества.
Суть вторично-активного транспорта можно раскрыть на примере переноса молекулы глюкозы, которая должна войти в клетку, где ее концентрация намного выше, чем во внешней среде. Следовательно, на ее перенос необходимо будет затратить энергию. Однако энергия тратится не собственно на перенос молекулы глюкозы, а на обратный транспорт иона натрия, в совокупности с которым она первоначально присоединяется к специфическому для натрия переносчику, и затем по натриевому градиенту концентрации проникает внутрь клетки. Внутри клетки глюкоза и натрий отщепляются от переносчика и глюкоза свободно по градиенту концентрации диффундирует через противоположную клеточную стенку, тогда как натрий выкачивается из клетки против градиента концентрации, т.е. с энергетическими затратами.
Эндоцитоз и экзоцитоз – это разновидности транспорта, при которых происходит изменение архитектуры мембраны, требующее энергетических затрат. Эндоцитоз – введение крупномолекулярных частиц из среды в клетку: фагоцитоз и пиноцитоз. Пиноцитоз в отличие от фагоцитоза сопровождается усвоением белковых молекул без предварительного их гидролиза. Например, новорожденные с молоком матери получают готовые антитела, способные к выполнению своих функций. Экзоцитоз – выделение крупных молекул из клетки (выделение квантов медиатора в синаптическую щель).
Электрическиеявленияввозбудимыхтканях
В основе процесса возбуждения лежит изменение потенциала покоя (ПП) или мембранного потенциала (МП), представляющего собой разность
Физиология. Конспект лекций |
-22- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Электрические явления в возбудимых тканях
электрических потенциалов внутри и снаружи клетки. МП всегда отрицателен и имеет постоянные значения для каждого типа клеток. Для нейронов потенциал покоя равен минус 70мВ, для мышечного волокна – минус 90 мВ. Отрицательные значения потенциала покоя связаны с тем, что в покоящейся клетке ионы натрия и калия постоянно перемещаются через клеточную мембрану. При этом К+ из клетки выходит в гораздо большем количестве, чем Na+ входит в клетку, т.к. проницаемость мембраны для ионов К+ примерно в 25 раз больше, чем для ионов Na+. Кроме того, органические анионы из-за своих больших размеров не могут выходить из клетки. В результате в состоянии покоя внутри клетки оказывается больше отрицательных, а снаружи
– больше положительных ионов. Возникающая разность потенциалов может быть описана уравнением Нернста:
ПП = 2,3RT/Flg[K+]нap./[K+]внутр.
Из уравнения следует, что, чем выше концентрация калия в среде, тем меньше величина этого соотношения и меньше соответственно величина МП, т.е. МП имеет калиевую природу. Однако расчетная величина МП всегда ниже, чем есть на самом деле. Например, по расчетам получается, что МП должен быть 90 мВ, а реально фиксируется – 70 мВ. Причина в том, что ионы натрия и хлора тоже вносят свой вклад в создание МП. В состоянии покоя открыта небольшая часть натриевых каналов (20-30 натриевых на 1000 калиевых), соответственно ионы натрия в небольших количествах могут поступать внутрь клетки. Для поддержания МП на постоянном уровне, т.е. для сохранения ионной асимметрии, служит натрий-калиевый и хлорный насос.
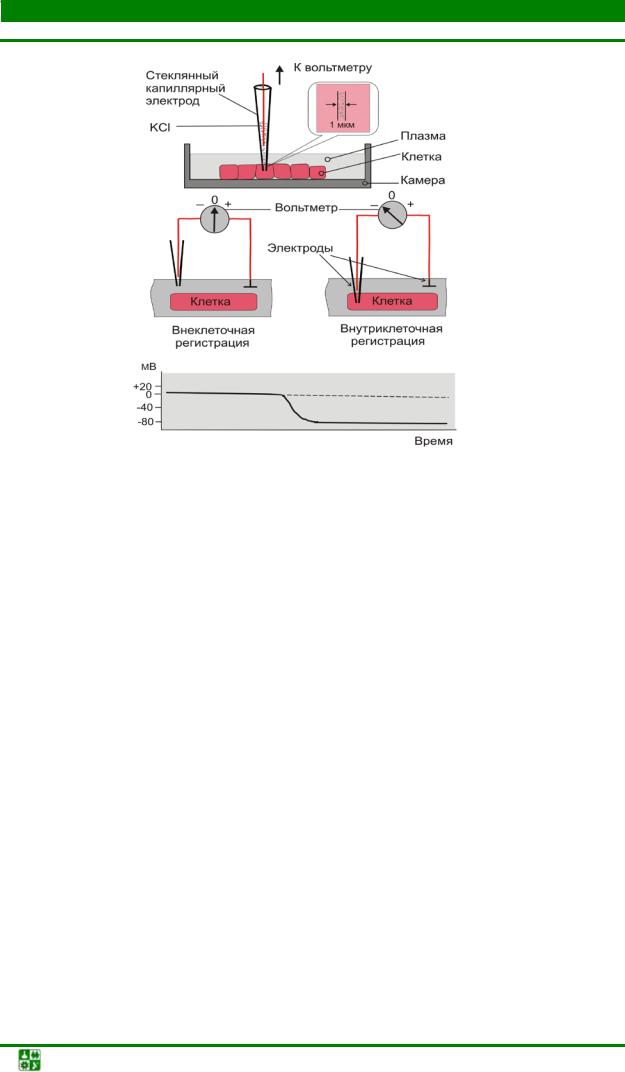
Регистрация МП осуществляется с помощью микроэлектродной техники (рис. 6). Клетка помещается в камеру, заполненную плазмой крови или физиологическим раствором. Датчиком потенциала является микроэлектрод, представляющий собой стеклянный капилляр с очень тонким кончиком (диаметром 0,5 мкм), заполненный электролитом (KCl).
Физиология. Конспект лекций |
-23- |
ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Электрические явления в возбудимых тканях
Рис. 6. Техника регистрации мембранного потенциала
Референтным электродом во внеклеточном пространстве служит хлорированная серебряная пластинка. Изначально оба электрода находятся во внеклеточном пространстве, и вольтметр регистрирует между ними нулевую разность потенциалов. Когда регистрирующий электрод вводят в клетку, то вольтметр показывает скачкообразный сдвиг потенциала в отрицательном направлении.
Несмотря на постоянную диффузию ионов МП клеток остается на одном уровне. Следовательно, кроме собственно ионных механизмов формирования мембранного потенциала, связанных с различной проницаемостью клеточной мембраны, имеет место активный механизм его поддержания. Таким механизмом являются ионные насосы, в частности натрий-калиевый насос.
Принцип работы натрий-калиевого насоса. Na/K-насос при каждом цикле переносит 3Na+ наружу и 2K+ внутрь клетки. Механизм переноса ионов Na+ и K+ через мембрану клетки можно представить следующим образом.
Внутри каналоподобной структуры насоса расположены активные центры (места связывания) Na+ и K+, которые поочередно вступают в контакт с внутри- и внеклеточной средой.
Физиология. Конспект лекций |
-24- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Электрические явления в возбудимых тканях
Рис. 7. Принцип работы натрий-калиевого насоса: А – связывание Na+ и отщепле-
ние K+, т.к. посадочные места в таком положении обладают большим сродством к Na+; Б - связывание молекулы АТФ и фосфорилирование фермента; В – выдвижение посадочных мест во внеклеточную среду; Г- освобождение ионов Na+ и связывание K+ (снижение сродства посадочных мест к Na+ и повышение – к K+); Д – дефосфорилирование АТФазы; Е – возврат системы в первоначальное состояние
Циклическое изменение конформации насоса происходит путем фосфорилирования и дефосфорилирования (присоединение и отщепление остатка фосфорной кислоты) его белковой молекулы (рис. 7). Этот процесс сопровождается изменением сродства мест связывания к соответствующим ионам. Места связывания, направленные внутрь клетки, имеют низкое сродство к ионам калия и высокое – к ионам натрия. Связывание трех ионов натрия приводит к изменению конформации белка и последующему связыванию АТФ и
фосфорилированию фермента. Фосфорилирование фермента приводит к дальнейшему изменению конформации, в результате чего места связывания оказываются в контакте с внеклеточной средой, теряют сродство к ионам натрия и приобретают таковое к ионам калия. Связывание калия вызывает дефосфорилирование фермента и возвращает насос в первоначальное положе-
ние, а ионы калия высвобождаются во внутриклеточное пространство. Потенциал действия. В основе возбуждения нервных и мышечных
клеток лежит повышение проницаемости мембраны для ионов натрия в результате открывания натриевых ионных каналов и появления трансмембранных токов, что приводит к быстрому изменению МП в положительном направлении – потенциалу действия (ПД). Таким образом, ПД в отличие от МП
Физиология. Конспект лекций |
-25- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Электрические явления в возбудимых тканях
имеет натриевую природу. Процесс возбуждения включает в себя генерацию ПД, его распространение и специфический ответ ткани на этот потенциал (сокращение, выделение секрета). Уровень потенциала, при котором деполяризация мембраны приводит к запуску ПД, называется порогом ПД и чаще всего составляет 50 мВ. Разница между МП и порогом ПД называется кри-
тическим уровнем деполяризации (КУД). Чем ниже (по абсолютной вели-
чине) порог, тем меньше критический уровень деполяризации и выше возбудимость нейрона.
Рис. 8. Фазы потенциала действия
Потенциал действия (ПД) – это высокоамплитудный, быстро распространяющийся по мембране сигнал, обеспечивающий передачу информации. При регистрации ПД наблюдается типичный пикообразный потенциал, в котором выделяют следующие фазы (рис. 8):
– фазу деполяризации, сопровождающуюся быстрым нарастанием МП от отрицательных значений до положительного пика – овершута (перелета), составляющего≈ +30 мВ. То есть это фаза, когда происходит перез а- рядка мембраны;
– фазу реполяризации, сопровождающуюся восстановлением исходного уровня МП. В ней выделяют фазу быстрой и медленной реполяризации. Фаза медленной реполяризации представлена следовыми потенциалами – следовой негативностью (гиперполяризацией) и следовой позитивностью (деполяризацией).
Физиология. Конспект лекций |
-26- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Электрические явления в возбудимых тканях
Проводимость плазматической мембраны для ионов натрия достигает максимума гораздо раньше, чем ПД достигает своего пика. Поэтому по достижении пика деполяризации, когда натриевые каналы уже закрыты, а K+ активно диффундирует из клетки, начинается процесс реполяризации. Нарастание проницаемости мембраны для ионов калия происходит гораздо медленнее, чем таковое отмечалось для ионов натрия. Своего максимума она достигает тогда, когда реполяризация уже наполовину завершена. В результате ускоряется вторая фаза реполяризации и возникает следовая гиперполяризация.
На фоне медленной фазы реполяризации (ее следовой негативности) происходит активация Na+/K+ насоса и возвращение потенциала мембраны в исходное состояние. Кроме Na+/K+ насоса регуляцию K+ в межклеточном пространстве осуществляют глиальные клетки, обладающие высокой проницаемостью для ионов калия.
При действии раздражителя недостаточной силы возникают пассивные деполяризационные изменения МП – электротонические потенциалы или локальный ответ, амплитуда которого зависит от силы раздражителя (рис. 8).
Рефрактерныепериоды
С инактивацией Na+-каналов связана очень важная характеристика возбудимых тканей – рефрактерность – невозбудимость. Закрывшиеся натриевые каналы не сразу восстанавливают свою способность к активации. В связи с этим в течение всей фазы деполяризации потенциала действия и частично фазы реполяризации клетка теряет способность к возбуждению, что именуется фазой абсолютной рефрактерности – состоянием полной невозбудимости
(рис. 9).
Физиология. Конспект лекций |
-27- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Рефрактерные периоды
Рис. 9. Рефрактерные периоды
Постепенно натриевые каналы выходят из состояния инактивации и возбудимость нервной клетки медленно восстанавливается. Этот период времени носит название относительной рефрактерности. Однако чтобы возбудить клетку в этот период необходимо, увеличить силу раздражения. Во время следовой гиперполяризации, когда потенциал мембраны опускается ниже уровня потенциала покоя и порог возбуждения соответственно увеличивается, имеет место пониженная возбудимость – субнормальность. В случае же следовой деполяризации, когда порог возбудимости понижается, наблюдается повышенная возбудимость – экзальтация или супернормаль-
ность.
Наличие периодов абсолютной рефрактерности ограничивает максимальную частоту генерирования ПД. В этой ситуации изменения внешних сигналов кодируются лишь изменениями частоты ПД (частотный код). Так как период абсолютной рефрактерности нервной клетки составляет около 1 мс, соответственно, частота их возбуждения (лабильность) может достигать 1000 импульсов в секунду. Однако большинство возбудимых клеток имеет максимальную частоту ПД около 500 импульсов в секунду.
Форма ПД и его длительность зависит от вида возбудимой ткани (рис. 10). ПД нерва составляет около 1 мс, скелетной мышцы – 10 мс, клеток миокарда – 200 мс. В одних случаях форма ПД пикообразная, в других – платообразная. Платообразная форма ПД клеток миокарда обеспечивает ритмичность сердечных сокращений.
Физиология. Конспект лекций |
-28- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Рефрактерные периоды
Рис. 10. Форма и длительность ПД в зависимости от вида возбудимой ткани
Законыраздражениявозбудимыхтканей. Распространение возбуждения
Для возбуждения ткани необходимо наличие внешнего по отношению к этой ткани раздражителя (за исключением тканей, обладающих автоматией). Такими раздражителями являются нервный импульс или выделение медиатора. В целом выделяют два типа раздражителей: адекватные и неадекватные. Адекватные раздражители способны в «малых дозах» вызвать раздражение. Как правило, это раздражители, к действию которых ткань приспособилась в процессе эволюции. Неадекватный раздражитель способен вызвать возбуждение, но при этом необходимо приложить большую силу, в результате чего ткань может быть повреждена.
Для того чтобы раздражитель вызвал возбуждение, он должен быть: достаточно сильным – закон силы; достаточно длительным – закон времени; достаточно быстро нарастать – закон градиента.
Закон силы – раздражитель по силе должен быть пороговым или выше порогового (рис. 11).
Физиология. Конспект лекций |
-29- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Законы раздражения возбудимых тканей. Распространение
Рис. 11. Закон силы
Порог – это минимальная сила раздражителя, способная вызвать возбуждение. Например, чтобы вызвать возбуждение клетки, у которой МП = – 90 мВ, а КУД = –60 мВ, пороговая сила должна составлять –30 мВ. Для одиночных структур (нейрона, нервного волокна) такая зависимость носит название правила «все или ничего». В соответствии с этим правилом амплитуда ПД не зависит от силы раздражителя. То есть, если сила раздражителя достигла порогового значения – критического уровня деполяризации, возникает ПД одной и той же амплитуды независимо от того, на сколько выше порогового значения сила раздражителя.
Если рассматривать ту или иную структуру в целом, например, нервный ствол, содержащий несколько нервных волокон (аксонов), то и в этом случае каждое волокно реагирует по принципу «все или ничего». Однако если регистрируется суммарная активность объекта, то его амплитуда в определенном диапазоне будет испытывать влияние силы раздражителя. Например, при внеклеточном отведении ПД нервного ствола, содержащего аксоны, КУД которых составляет 1-й – 30 мВ, 2-й, 3-й и 4-й – 40 мВ, 5-й,6-й,7-й и 8-й
– 50 мВ. При воздействии с силой в –30 мВ будет возбуждаться 1 аксон, при
–40 мВ – 4аксона, а при –50 мВ – 8 аксонов. То есть в пределах от –30 до –50 мВ амплитуда регистрируемого ПД будет возрастать, тогда как при дальнейшем увеличении силы раздражителя амплитуда будет оставаться постоянной. Самым возбудимым нервным волокном в данном случае, является 1-й аксон.
Закон времени утверждает, что раздражитель, вызывающий возбуждение, должен быть достаточно длительным. При этом чем меньше по времени действует на ткань раздражитель, тем больше должна быть его сила. Наглядно этот закон может быть продемонстрирован с помощью кривой (гиперболы) Гоорвега-Вейса-Лапика, из которой следует, что если раздражитель
Физиология. Конспект лекций |
-30- |
ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Законы раздражения возбудимых тканей. Распространение
достаточно длительный, то пороговая сила раздражителя уже не зависит от длительности (рис. 12). Эта минимальная (пороговая) сила называется реоба-
зой.
Минимальное время, в течение которого раздражитель данной силы должен воздействовать на ткань, обозначается термином «полезное время». Если сила раздражителя равна двум реобазам, то полезное время такого раздражителя называют хронаксией. Хронаксия широко применяется в клинической практике при лечении ранений нервов.
Рис. 12. Кривая силы и времени (Гоорвега-Вейса-Лапика)
Поражение нерва приводит к увеличению хронаксии, т.е. для его возбуждения требуется приложить большую силу, чем к такому же здоровому.
Закон градиента также носит гиперболический характер. Чтобы раздражитель вызвал возбуждение, он должен достаточно быстро нарастать. Медленное нарастание приводит к повышению порога раздражения, что соответственно требует воздействия большей силы. Минимальная скорость нарастания раздражителя, при которой ткань еще способна ответить возбуждением, называется минимальным градиентом. Минимальный градиент той или иной структуры тем выше, чем выше скорость проведения импульса. Например, минимальный градиент нерва выше такового скелетной мышцы.
Распространение возбуждения. Протекающие по принципу «все или ничего» процессы возбуждения в отдельных участках мембраны сопряжены друг с другом посредством электротонического распространения токов вдоль волокна (рис. 13). Принцип распространения ПД связан с возникновением входящих локальных токов ионов натрия через возбужденный участок мем-
Физиология. Конспект лекций |
-31- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Законы раздражения возбудимых тканей. Распространение
браны, которые приводят к возбуждению соседнего неактивного участка (рис. 14). При этом в соседних неактивных участках нервного или мышечного волокна возникают локальные выходящие токи, вызывающие перераспределение зарядов на мембране и деполяризацию уже в этих участках.
Рис. 13. Возникновение местных электрических токов
Рис. 14. Проведение потенциала действия в безмиелиновых (А) и миелиновых нервных (Б) волокнах
Как только деполяризация в этих участках достигает порогового уровня, в них открываются натриевые каналы и возникает ПД. Другими словами, проведение ПД в немиелинизированных волокнах связано с его постоянным возникновением в соседних участках мембраны. Распространение возбуждения происходит без снижения скорости и амплитуды ПД.
Скорость проведения ПД зависит от того, насколько быстро и насколь-
Физиология. Конспект лекций |
-32- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Законы раздражения возбудимых тканей. Распространение
ко далеко от активного участка происходит деполяризация мембраны до порогового уровня при протекании локальных токов. Это в свою очередь зависит от величины входящего ионного тока и кабельных свойств волокна. Величина входящего тока определяется плотностью натриевых каналов мембраны, а кабельные свойства – удельным сопротивлением мембраны и аксоплазмы, а также диаметром волокна. Чем толще нервное волокно, тем дальше будет распространяться деполяризация от активного участка, соответственно, выше будет скорость распространения ПД.
Для того чтобы увеличить скорость распространения ПД в относительно тонких нервных волокнах, природа создала на них специальную миелиновую оболочку, образованную мембранами глиальных швановских клеток. Миелин обладает высоким сопротивлением, поэтому ток не течет в межперехватных участках. Только короткие участки миелинизированных волокон – перехваты Ранвье – имеют обычную клеточную мембрану, которая в отличие от мембраны безмякотных волокон имеет очень высокую (в 100 раз большую) плотность натриевых каналов. В таких волокнах возбуждение передается сальтаторно, перескакивая с одного перехвата на другой. Задержка проведения возбуждения в этом случае происходит только в местах перехватов, где электротонический потенциал должен достигнуть порога и вызвать ПД.
Контрольныевопросыизадания
1.Возбудимые ткани и их свойства.
2.Типы транспорта веществ через биологические мембраны.
3.Какова природа мембранного потенциала?
4.Произойдет ли изменение мембранного потенциала при действии раздражителя, составляющего 95 % от порогового?
5.Природа и механизм развития потенциала действия.
6.Принцип работы натрий-калиевого насоса.
7.Законы раздражения возбудимых тканей.
8.Распространение возбуждения по миелинизированным и немиелинизированным волокнам.
9.Что произойдет с мембранным потенциалом, если внутри аксона повысить концентрацию ионов калия?
10.Рассчитайте лабильность для нервных волокон группы А, если известно, что длительность потенциала действия = 0,5 мс.
Физиология. Конспект лекций |
-33- |

ЛЕКЦИЯ 2. ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Контрольные вопросы и задания
11.Какая ткань обладает большей возбудимостью: а) критический уровень деполяризации (КУД) которой равен –60 мВ; б) КУД = – 50 мВ, если мембранный потенциал равен –80 мВ?
12.Как изменится мембранный потенциал, если заблокировать работу Na/K- зависимой АТФ-азы?
Физиология. Конспект лекций |
-34- |