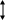Гомельский государственный медицинский институт
Кафедра микробиологии и вирусологии
Лекция
Тема: "Онкогенные вирусы"
Подготовил: Бондарева М.Н.
Онковирусы – это вирусы, обладающие способностью обуславливать трансформацию нормальных клеток эукариот в опухолевые.
В 1903 г. А. Боррель и Боск впервые высказали гипотезу о вирусной природе агентов вызывающих раковые опухоли. В 1968 г. Л.А. Зильбером предложил вирусо-генетическую теорию возникновения опухолей.
Классификация онковирусов:
РНК-содержащие онковирусывирусы
Семейство: Retroviridae
Подсемейство: Oncovirinae
Саркомно-лейкозный комплекс птиц
Саркомно-лейкозный комплекс мышей
Вирус рака молочных желез мышей, крыс и обезьян
Вирусы, вызывающие лейкозы и саркомы у крыс, хомяков, кошек.
ДНК-содержащие онковирусывирусы
Семейство: Papovaviridae
Род: Papillomavirus
Вирусы, вызывающие папилломы у кроликов, человека, собак
Род: Poliomavirus
Вирус палиомы типа 1, вызывающий саркому у крыс, мышей и хомячков.
Вирус палиомы типа 2 ОВ-40.
Семейство: Adenoviridae
Род: Adenovirus
Аденовирусы человека, обезьян, птиц.
Семейство: Poxviridae
Род: Lepopoxvirus
Вирусы группы оспы, вызывающие фиброму у кроликов, белок
Вирус Яба
Вирус контагиозного моллюска человека
Семейство: Herpesviridae
Герпесвирус человека, болезни Марика кур, лимфомы и саркомы обезьян
Нуклеиновые кислоты этих вирусов могут иметь линейные и концевые формы молекул с повторами на концах, иногда с перекрывающими друг друга концами.
Строение РНК-содержащих вирусов
Онкорновирусы являются естественными возбудителями большинства злокачественных образований животных с установленной вирусной природой. Это вирусы со сложным, полиморфным типом симетрии диаметром около 100 нм, имеющих единый план строения.
Вставить картинку и стереть эту гадкую надпись
Структура онкорновирусов:
Однонитчатая нуклеиновая кислота
Структурные и неструктурные белки (ферменты)
Двойная мембрана (наружная и внутренняя)
Липопротеиновая оболочка (защита от клеточных нуклеаз)
Антигенные детерминанты: Gs – антиген (основной белок нуклеотида) группоспецифический антиген; типоспецифические антигены – белки оболочки в виде поверхностных выступов.
По морфологии онкорновирусы делятся на типы:
А, являющийся внутриклеточным предшественником онкорновирусов типа B, C, D.
Б, являющийся возбудителем карцином молочных желез у мышей.
С, являющийся возбудителем лейкозов и саркомы птиц и млекопитающих
D, объединяет вирус перевиваемых раковых клеток человека, эндогенные вирусы, белок и обезьян.
Структура и морфогенез ДНК-содержащих онковирусов сходны с организацией и внутриклеточным развитием инфекционных вирусов оспы, герпеса, аденовирусов и папововирусов.
Репликационный цикл днк-содержащих вирусов в перемессивной системе
Возможно два пути реализации генетической информации ДНК-содержащих онковирусов. репликационный цикл, характерный для ДНК содержащих вирусов; интеграция генетической информации ДНК-содержащих онковирусов в геном клетки хозяина с последующей ее репликацией в интегрированном состоянии. Каждый онковирус имеет свою ферментную систему или один ключевой фермент который может осуществлять, с одной стороны, генетическую рекомбинацию (интеграцию вирусной ДНК), а с другой – в пермиссивных условиях обеспечивать репликацию вирусной ДНК Основные этапы этого процесса:
Осуществляются разрывы родительских ДНК на некотором расстоянии друг от друга, с предварительным расщеплением двойных спиралей взаимодействующих молекул ДНК, (разрываться должны полинуклеотидные нити клеточной и вирусной ДНК), причем разрыв в одной комплиментарной нити должен быть несимметричен разрыву во второй. Образуются структуры с однонитчатыми комплиментарными ("липкими") концами. В результате у каждого свободного конца нити появляется возможность присоединять чужой конец нити ДНК по принципу комплиментарности (при условии гомологии нуклеотидных последовательностей), образуется двойная спираль нового генома. Встраивание вирусной ДНК в хромосому клетки осуществляется вирусспецифической эндонуклеазой, которая производит разрывы в полинуклеотидных цепях ДНК вируса как вируса, так и клетки.
При продуктивной инфекции эндонуклеаза производит разрывы одной нити суперциркулярной молекулы ДНК онковирусов в одной определенной точке, являясь тем самым индуктором ее синтеза. При абортивной инфекции этот фермент вызывает одиночные разрывы во определенных точках вирусных и клеточных молекул ДНК, обеспечивая рекомбинацию. В трансформированных клетках он отсекает вирусную ДНК от клеточной.
В процессе рекомбинации ДНК вновь синтезированные участки представляют собой отдельные фрагменты, которые затем сшиваются ДНК-лигазой, при этом может происходить объединение вирусных и клеточных участков ДНК. Таким образом встраивание ДНК вируса осуществляется аналогично интеграции умеренного фага.
Этапы трансформации, индуцированной рнк-содержащими онкогенными вирусами
В вирионах онкорнавирусов содержатся все ферменты, необходимые для репродукции вируса, синтеза вирусной ДНК, трансляции информации с РНК на ДНК и для встраивания ДНК-провируса в хромосому клетки (фермент, осуществляющий рекомбинацию и репарацию ДНК). Онкорнавирусы содержат в своем составе РНК-зависимую ДНК полимеразу, для которой матрицами являются односпиральные РНК, двуспиральные РНК и ДНК, а также ДНК-РНК-гибриды; эндонуклеазу ДНК, разрывающую фосфодиэфирные связи внутри остова молекулы.
Исходя из специфики онкорнавирусов (каждый из них имеет свой ДНК-провирус), говорят о существовании двух их форм: соматической и зародышевой.
Соматическая форма онкорнавирусов.
Представляет собой экзогенную вирусную ДНК, т.е. синтезированную в процессе экзогенной инфекции онкорнавирусами. Механизм включения этих ДНК в клеточный геном идентичен встраиванию молекул ДНК-фагов или ДНК-содержащих онковирусов в хромосому клетки:
РНК-зависимый синтез второй нити ДНК на матрице вирусной РНК.
ДНК-зависимый синтез второй нити ДНК на нити ДНК промежуточного гибридного продукта
Оба этапа осуществляются с помощью обратной транскриптазы (ревертазы). Такой синтез ранней ДНК требует постоянного функционирования этой ДНК на протяжении всего времени инфекции, т.к. она является матрицей для синтеза вирусной РНК, которая, как и родительские РНК обеспечивают синтез структурных белков вирионов.
Начальный синтез ДНК происходит в репликационном комплексе, содержащем молекулы ДНК, связанные водородными связями с молекулами вирусной РНК. Каждая молекула ДНК вытесняется как однонитевая молекула в результате синтеза последующей нити ДНК. Затем следует синтез двуспиральной ДНК на матрице односпиральной ДНК (до или после того, как она сойдет с РНК-матрицы).
ДНК двунитчатая, репликативная форма
РНК вирусная однонитчатая
Интеграция с клеточной ДНК
ДНК


Биологическое значение двуспиральной вирусной ДНК:
интеграция с комплиментарными участками клеточной ДНК и модификации генов клетки и вируса.
может функционировать в интегрированном состоянии матрицей для синтеза вирусных и РНК (непрерывный источник информации для поддержания неопластического типа клеток.
интегрированная ДНК может защищать генетическую информацию вирусной РНК от действия нуклеаз и тем самым выполнять функцию репликации вируса.
Весь описанный комплекс макромолекулярных синтезов заканчивается интеграцией вирусной ДНК с клеточной.
Гены онкорнавирусов могут длительное время сохраняться в геноме зараженных и трансформированных клеток.
Существует 3 типа экспрессии вирусного генома, включенного в хромосомы и начавшего функционировать и реплицироваться вместе с генами хозяина.
1 тип: Полная транскрипция и трансляция вирусного генома, обуславливающая спонтанную продукцию инфекционного вируса.
2 тип: частичная экспрессия вирусной информации, которая, например, может проявляться в синтезе только Gs антигенов.
3 тип: связан с полной репрессией вирусного генома.
Репликация вирусного генома происходит в течение первых часов после заражения, а трансформации подвергаются обе дочерние клетки, происходящие из инфицированной материнской клетки.
Зародышевая форма онкорнавирусов.
Зародышевый ДНК-провирус онконавирусов состоит из нескольких генов, в которых закодирована информация на синтез трансформирующих белков, продукцию Gs антигенов, типоспецифических белков, обратной транскриптазы и других ферментов. В нормальных клетках онкоген, как и вироген, находится в реплицированном состоянии, а их экспрессия имеет место в основном в эмбриональных и недифференцированных, пролиферирующих клетках. В зрелых, непролиферирующих клетках экспрессия. ДНК-провируса приводит к опухолевому прерождениюклетки.
Схема виро- и онкогена зародышевого ДНК-провируса.
Экспессия этих "раковых" генов может происходить вследствие мутации генов-регуляторов, контролирующих провирус, под воздействием химических, физических канцерогенов, гормонов, старения и т.д.
Выделенные две формы ДНК-провирусов не исключают взаимно друг друга, а в естественных условиях существует переход одной формы в другую.
Две формы существования онкорнавирусов.
Соматический ДНК-провирус


Интеграция с ДНК клетки – зародышевый
ДНК провирус

Основное звено канцерогенеза – фиксация в клеточном геноме информации о неопластическом преобразовании клетки с помощью ревертаз; депрессия или мутация информации онковирусов (ДНК геномов ДНК содержащих вирусов, или ДНК-копий онкорнавирусов.
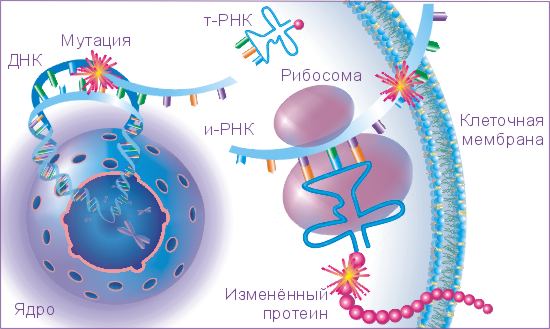
ОСНОВНОЕ ОТЛИЧИЕ В РЕПЛИКАЦИОННОМ ЦИКЛЕ ОНКОРНАВИРУСОВ И ДНК- СОДЕРЖАЩИХ ОНКОГЕННЫХ ВИРУСОВ:
Инфекция ДНК-содержащими онковирусами
Инфекция РНК-содержащими онковирусами
Репликация
1) Интеграция
Интеграция
2) Репликация



и и
У онкорнавирусов процесс интеграции является одним из этапов цикла в пермиссивной системе. Так образовавшиеся вирусные частицы отпочковываются от клеточной мембраны, после чего проникают в другие клетки, где с помощью ревертазы транскрибируются в ДНК. Вновь синтезированные молекулы вирусной ДНК переносятся в ядро и пройдя предварительную стадию циклизации, интегрируются в геноме клетки.
Опухолевая трансформация клеток представляет сложный макромолекулярно-генетический процесс, который обуславливается как вирусными, так и клеточными ферментными системами, но инициатором и регулятором неопластической трансформации является вирус, (продукты его онкогенов)
На первом этапе трансформации в основном осуществляется функциональный генный дисбаланс, т.е. происходит наследственно закрепляющаяся перестройка работы нового генома трансформируемой клетки (утрата и приобретение новых копий генов, в следствие чего может возрастать активность отдельных изоферментов.
Генетическая рекомбинация,
осуществляемая вирускодированной
эндонуклеазой
III Зародышевый ДНК-провирус
(ДНК-транскрит онкорнавирусов или геном
ДНК-содержащего онковируса, передающийся
через зародышевую клетку потомству)
Стабильная трансформация
Индуцированная рекомбинация (разрыв
хромосом рентгеновским излучением,
химическими канцерогенами и т.д.,
способствующие встраиванию вирусного
генома в клеточный).
I Абортивная трансформация
(присутствие не фиксированного, частично
или полностью активного вирусного
генома в клетке).
II Стабильная трансформация
соматических клеток, обусловленная
включением вирусного генома ДНК-содержащих
онковирусов и соматического ДНК-провируса
онкорнавирусов в хромосому клетки-хозяина.
Снятие блока с раковых генов
ДНК-провируса (УФ-, рентгеновское и
гамма-излучение, химические канцерогены,
лизосомальные ДНК-азы, возрастные
факторы и т.д.)