

Физиология. Курс лекций
.pdf31
каналы, по которым кальций входит в межфибриллярное пространство миофибриллы, что приводит к инициации сокращения. Потом мембраны продольных трубочек СР начинают активно откачивать ионы кальция назад в СР, и его концентрация в межфибриллярном пространстве уменьшается, что приводит к расслаблению.
Рис.5. Поперечный срез миофибриллы дает представление о гексагональном распределении актиновых и миозиновых нитей.
Z — линии, разделяющие два соседних саркомера; J — изотропный диск;
А — анизотропный диск; Н — участок с уменьшенной анизотропностью.
32
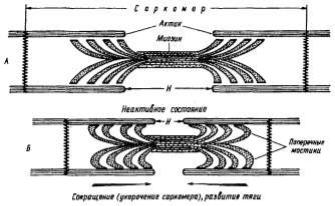
Рис.6. Сокращение мышцы.
А. Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии. Б. Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги.
Поскольку возврат ионов кальция в цистерны саркоплазматического ретикулума идет против диффузионного градиента, то этот процесс требует затрат энергии. Ее источником служит АТФ. Одна молекула АТФ затрачивается на возврат 2-х ионов кальция из межфибриллярного пространства в цистерны. При снижении содержания ионов кальция до подпорогового уровня (ниже 10-8м) молекулы тропонина принимают форму, характерную для состояния покоя. При этом вновь тропомиозин блокирует участки для прикрепления поперечных мостиков к нитям актина. Все это приводит к расслаблению мышцы вплоть до момента прихода очередного потока нервных импульсов, когда описанный выше процесс повторяется. Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения.

33
Рис.7. Схема элементов нервно-мышечного синапса. 1 — миэлиновая оболочка аксона; 2 — концевые веточки аксона;
3 — пузырьки, содержащие ацетилхолин;
4 — митохондрия;
5 — пресинаптическая мембрана, покрывающая концевую веточку аксона в зоне нервно-мышечного синапса;
6 — синаптическая щель;
7 —постсинаптическая мембрана, покрывающая мышечное волокно в зоне нервно-мышечного синапса;
8 — ацетилхолинорецепторы на постсинаптической мембране;
9 — митохондрия мышечного волокна;
10 — ядро мышечной клетки;
11 — миофибрилла.
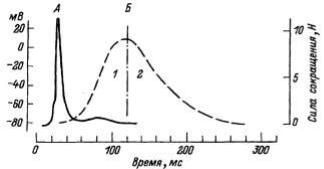
Рис.8. Схема временной последовательности развития потенциала действия (ПД), освобождения ионов кальция (Са2+) и развития изометрического сокращения мышцы.
34
Режимы и типы мышечных сокращений. Режим сокращения мышечных волокон определяется частотой импульсации мотонейронов. Механический ответ мышечного волокна или отдельной мышцы на однократное их раздражение называется одиночным сокращением. При одиночном сокращении выделяют: 1. фазу развития напряжения или укорочения; 2. фазу расслабления или удлинения (рис.9). Фаза расслабления продолжается примерно в два раза дольше, чем фаза напряжения. Длительность этих фаз зависит от морфофункциональных свойств мышечного волокна: у наиболее быстро сокращающихся волокон глазных мышц фаза напряжения составляет 7-10 мс, а у наиболее медленных волокон камбаловидной мышцы — 50100 мс.
Рис.9. Развитие во времени потенциала действия (А) и изометрического сокращения мышцы, приводящей большой палец кисти (Б).
1 — фаза развития напряжения; 2 — фаза расслабления.
Вестественных условиях мышечные волокна двигательной единицы и скелетная мышца в целом работают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон. Так, режим одиночного сокращения медленных волокон камбаловидной мышцы человека обеспечивается при частоте импульсации мотонейрона менее 10 имп/с, а быстрых волокон глазодвигательных мышц — при частоте импульсации мотонейрона менее 50 имп/с.
Врежиме одиночного сокращения мышца способна работать длительное время без развития утомления. Однако в связи с тем, что длительность одиночного сокращения невелика, развиваемое мышечными волокнами напряжение не достигает максимально возмож-

35
ных величин. При относительно высокой частоте импульсации мотонейронов каждый последующий раздражающий импульс приходится на фазу предшествующего напряжения волокона, то есть до того момента, когда оно начинает расслабляться. В этом случае механические эффекты каждого предыдущего сокращения суммируются с последующим. Причем величина механического ответа на каждый последующий импульс меньше, чем на предыдущий. После нескольких первых импульсов последующие ответы мышечных волокон не изменяют достигнутого напряжения, а лишь поддерживают его. Такой режим сокращения называется гладким тетанусом (рис.10-11). В подобном режиме двигательные единицы мышц человека работают при развитии максимальных изометрических усилий. При гладком тетанусе развиваемое ДЕ напряжение в 2-4 раза больше, чем при одиночных сокращениях.
Аксон разветвляется на небольшое число концевых веточек и иннервирует небольшую группу мышечных волокон. У мотонейронов медленных ДЕ сравнительно низкая частота разрядов (6-10 имп/с). Они начинают функционировать уже при малых мышечных усилиях. Так, мотонейроны ДЕ камбаловидной мышцы человека при удобном стоянии работают с частотой 4 имп/с. Устойчивая частота их импульсации составляет 6-8 имп/с. С повышением силы сокращения мышцы частота разрядов мотонейронов медленных ДЕ повышается незначительно. Мотонейроны медленных ДЕ способны поддерживать постоянную частоту разрядов в течение десятков минут.
Рис.10. Одиночные (а) и тетанические (б,в,г,д) сокращения скелетной мышцы. Накладывание волн сокращения друг на друга и образование тетануса при частотах раздражения: 6—15 раз в 1 с; в — 20 раз в 1 с; г — 25 раз в 1 с; д — более 40 раз в 1 с (гладкий тетанус).

36
Рис.11. Двигательные единицы (ДЕ) мышцы и их типы.
1— медленные, слабые, неутомляемые мышечные волокна. Низкий порог активации мотонейрона;
2— промежуточный тип ДЕ;
3— быстрые, сильные, быстроутомляемые мышечные волокна. Высокий порог активаций мотонейрона.
Прирост силы при изометрическом сокращении суммируется с пассивным напряжением мышцы. Пиковые (максимальные) напряжения в этих условиях называют максимум изометрического напряжения (рис.12). Напряжение сокращающейся мышцы максимально, если ее длина составляет примерно 120 % от равновесной. Это состояние носит название длины покоя.
Укорочение мышцы меньше длины покоя или ее растяжение больше этой величины приводит к снижению силы сокращения.

37
Рис.12. Зависимость между длиной мышцы и силой сокращения (А); между скоростью укорочения и величиной внешней нагрузки (Б).
3. Виды сокращения мышц: одиночное, тетаническое, изотоническое, изометрическое, тонус мышц
Возбуждение и сокращение мышц при естественных двигательных актах вызывается нервными импульсами, поступающими из центральной нервной системы.
Раздражение двигательного нерва, вызывающее сокращение мышцы, называют непрямым раздражением, а непосредственное раздражение самой мышцы — прямым раздражением. Поскольку возбудимость мышечных волокон ниже, чем нервных, для получения истинного прямого раздражения необходимо выключить в мышце двигательные нервные окончания или ввести микроэлектроды в мышечные волокна.
Запись сокращений мышц на приборе миографе при прямом или непрямом раздражений называется миографией. Отведение биопотенциалов самой мышцы при ее возбуждении (с помощью

38
приложенных или вколотых электродов) называется электромиографией. Ее используют для записи электрической активности не только скелетных мышц, но и гладких мышц внутренних органов. Скорость и сила ответной реакции скелетной мышцы на раздражение зависит не только от параметров раздражителя, но и от динамических свойств составляющих ее волокон.
По скорости сокращения различают два типа мышечных волокон, а точнее, два типа двигательных единиц — быстрые и медленные. Что касается самих мышц, то правильнее говорить о смешанных типах мышц — с преобладанием быстрых или медленных волокон. В быстрых, физических, волокнах обычно лучше развит саркоплазматический ретикулум, что способствует быстрому выбросу ионов Са++. Они слабее снабжены кровеносными сосудами, имеют более крупные и длинные волокна, обладают и другими особенностями (см. схему). Их расслабление после сокращения происходит в 50—100 раз быстрее, чем медленных волокон.
Режимы сокращения мышцы определяются частотой и силой стимуляции. Сокращение наступает не сразу после нанесения раздражения, а через определенный промежуток времени, называемый латентным периодом. Его величина составляет для икроножной мышцы лягушки 0,01 с. Фаза укорочения длится 0,04 с, фаза расслабления — 0,05 с (рис.13). Начало сокращения соответствует восходящей фазе потенциала действия, когда он достигает пороговой величины (примерно 40 мВ).
Рис.13. Сокращение скелетной мышцы:
А— одиночное сокращение (окрашенный участок — латентный период);
Б— феномен «лестницы» (увеличение амплитуды первых сокращений).
При нанесении на мышцу повторных раздражений постоянной силы, длительности и частоты (ниже тетанизирующей) наблюдается «феномен лестницы», т. е. увеличения амплитуды сокращений на
39
первые несколько раздражений (рис.13Б). Вероятно, это обусловлено постепенным увеличением количества ионов Са+ +, связываемых тропонином.
Сокращение каждого отдельного быстрого мышечного волокна при одиночных раздражениях подчиняется закону «все или ничего», т. е. генерация распространяющегося потенциала действия, полное высвобождение ионов Са++ и максимальное по силе сокращение миофибрилл происходит при определенной пороговой интенсивности стимула. Однако при электрическом раздражении целой мышцы сила сокращения зависит от интенсивности стимула. Это обусловлено тем, что в состав мышцы входят волокна разной толщины и разной возбудимости. Амплитуда сокращения мышцы будет возрастать до тех пор, пока сила раздражения не станет пороговой для всех волокон, входящих в состав мышцы.
Если на мышцу действуют два быстро следующих друг за другом раздражения (период между стимулами не более 100 мс), мышечные волокна расслабляются не полностью и каждое последующее сокращение как бы наслаивается на предыдущее. Это связано с тем, что кальциевый насос не успевает возвращать все ионы Са++ в саркоплазматический ретикулум в промежутках между стимулами. Происходит суммация (суперпозиция) сокращений, которая может быть полной, когда оба сокращения сливаются, образуя одну вершину, или неполной, в зависимости от частоты раздражений. В обоих случаях сокращение будет иметь большую амплитуду, чем максимальное сокращение при одиночном раздражении.
При воздействии на мышцу ритмических раздражений высокой частоты наступает сильное и длительное сокращение мышцы. Такое состояние устойчивого сокращения называется тетанусом. Он может быть зубчатым (при частоте раздражения 20—40 Гц) или сплошным, гладким (при частоте 50 Гц и выше). Амплитуда тетанического сокращения (напряжения) в 2— 4 раза выше амплитуды одиночного сокращения при той же силе раздражения. Минимальный промежуток времени между отдельными эффективными стимулами при этом не может быть меньше, чем рефрактерный период, соответствующий приблизительно длительности потенциала действия (3 мс).
В естественных условиях мышечной деятельности мотонейроны посылают к мышцам не одиночные импульсы, а серии импульсов. Частота их в разных условиях и в разных двигательных единицах
40
может быть различной (рис.14). При относительно низкой частоте импульсации мышечные волокна работают в режиме одиночных сокращений (для медленных двигательных единиц эта частота реже, чем для быстрых). При относительно высокой частоте импульсации мотонейронов возникает тетанический режим работы мышечных волокон. Эта частота также не одинакова для быстрых и медленных волокон.
При тетанических сокращениях мышечные волокна утомляются больше, чем при одиночных сокращениях. Поэтому даже в пределах одной мышцы происходит периодическая смена частоты импульсации (вплоть до полного исчезновения) в разных двигательных единицах.
Импульсы с мотонейронов в условиях покоя участвуют в поддержании так называемого мышечного тонуса. Под тонусом понимают состояние естественного постоянного напряжения мышц при невысоких энергетических затратах. Это состояние регистрируется электромиографически. Пассивный мышечный тонус зависит от вязкоэластичных свойств мышц, а активный мышечный тонус поддерживается асинхронной работой части медленных нейрофункциональных единиц. В поддержании тонуса участвуют проприо-рецепторы мышц (мышечные веретена) и центральная нервная система. Типы сокращений. При сближении актиновых и миозиновых фибрилл вследствие замыкания поперечных мостиков в мышечном волокне развивается напряжение (активная механическая тяга). Реализуется оно по-разному в зависимости от условий, в которых происходит сокращение мышц, прежде всего от величины внешней нагрузки. Различают два основных типа мышечных сокращений — изотонический и изометрический.
Изотоническое (от греч. isos — равный, tonos — напряжение) — это такое сокращение, при котором происходит укорочение мышечных волокон, но их напряжение не меняется. В эксперименте изотоническое сокращение получают при электрическом (тетаническом) раздражении изолированной мышцы, отягощенной небольшим грузом (рис.15). Укорочение мышцы происходит при постоянном напряжении, равном внешней нагрузке.
Изометрическое (от греч. isos — равный, meros — мера) — это сокращение, при котором длина волокон не уменьшается, но их напряжение возрастает.