

Регуляция лимфопоэза.
В-лимфопоэз. Как и в случае миелоидных линий, развитие В-клеток начинается с дифференцировки полипотентной стволовой клетки в недифференцированные, но коммитированне предшественники В-клеток. Пролиферация и дифференцировка этих предшественников В-клеток на начальных стадиях индуцируется интерлейкином-7 (ИЛ-7) и фактором стволовых клеток (ФСК). Как только образовались распознаваемые пре-В- и В-клетки, происходит их дальнейшая дифференцировка и деление при участии иммуноглобулинового антигенного рецептора, стимулируемая растворимым ИЛ-4 и ИЛ-6.После формирования плазматических клеток, производящих АТ, ИЛ-6 и КСФ-ГМ стимулируют дополнительную пролиферацию и секрецию АТ
Т-лимфопоэз. Первоначально пре-Т-клетки подвергаются сложной негативной и позитивной селекции в тимусе, в процессе которой они "обучаются" распознавать "свои" и "не свои" (чужеродные) клетки. Возникающие в результате зрелые Т-клетки являются субъектами антиген- и цитокинининдуцированной активации и экспансии. Стимуляция АГ-ном в присутствии ИЛ-2 или АГ-ном в сочетании со стимуляцией вспомогательными макрофагами либо дендритными клетками, ведет к прямой активации Т-клеток.
В регуляции лимфопоэза принимают участие также интерфероны. Их три типа - α, β и γ. Их синтез осуществляется в основном моноцитарными клетками и активированными Т-лимфоцитами. Действие интерферонов на лимфоидные клетки достаточно сложно. В малых дозах они могут оказывать стимулирующее действие на пролиферацию и дифференцировку лимфоцитов, в то время как большие дозы подавляют эти процессы.
Эритроцитарный росток.
4 класс - эритробласт - первая морфологически определимая клетка красного ростка. Ядро круглое, занимает большую часть клетки, имеет нежную сетчатую структуру и 2-4 ядрышка. Цитоплазма ярко-базофильная без просветлений вокруг ядра, в виде узкого ободка окружает ядро. По мере созревания ЭКЦ, начиная с ЭБ, размеры ядра и клетки в целом уменьшаются.
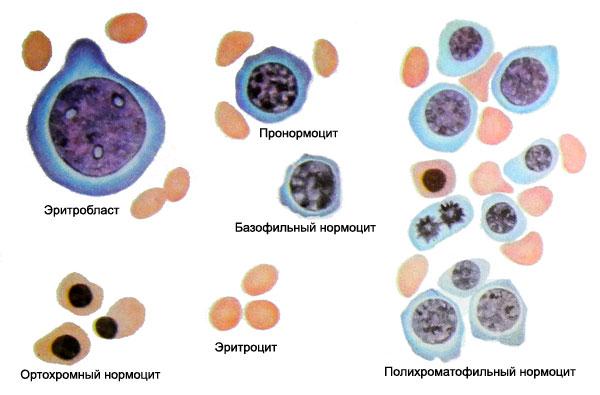
5 класс - пронормоцит (проэритробласт) напоминает эритробласт, но в отличие от него меньшего диаметра, имеет резко очерченное округлое ядро, структура хроматина более грубая, местами утолщена, более компактна, отсутствуют нуклеолы. Цитоплазма резко базофильная с легким перинуклеарным просветлением.
Нормоциты. В зависимости от насыщения Hb-ном они делятся на базофильные, полихроматофильные, оксифильные. Hb накапливается в цитоплазме при участии ядра. Поэтому первые скопления Hb наблюдаются в перинуклеарной зоне. Базофильные нормоцит имеет ядро округлой формы, с грубой структурой хроматина с чередованием темных и светлых участков, напоминая колесо со спицами. Цитоплазма базофильная.
Полихроматофилы отличаются от базофильного окраской цитоплазмы, воспринимают и кислые и основные красители (на рисунке КМ и кровь).
Оксифильные по величине приближаются к эритроциту, имеют небольшое пикнотичное ядро, оксифильную цитоплазму. Это полностью насыщенная гемоглобином клетка, цитоплазма окрашена в розовый тон, воспринимает кислые краски( на рисунке двухядерный плазмоцит, справа оксифильный нормоцит).
С началом гемоглобинизации цитоплазмы происходит инволюция ядра, пикноз. Ядро приобретает грубую радиарную (колесовидную) структуру, затем становится грубым, пикнотическим ("вишневая косточка"). Клетка освобождается от ядра путем кариорексиса, реже кариолизиса.
6 класс - ретикулоцит. В нормальных условиях часть полихроматофильных нормоцитов теряют ядро, превращаясь в ретикулоцит. Он является незрелым эритроцитом. В нем определяется сетчатая субстанция, которая является артефактом и состоит из агрегированных митохондрий, комплекса Гольджи, рибосом и других органелл, которая при прижизненной окраске (бриллиантовым крезиловым синим, акридиновым оранжевым) выявляется в виде сетчатой субстанции. В мазках, окрашенных обычным способом, сетчатая субстанция не видна, и эритроциты имеют полихроматофильную окраску.
Образовавшийся в КМ ретикулоцит пребывает в нем 36-44 часа, а затем попадает в кровь, где дозревает в течение 24-30 часов. В период пребывания ретикулоцитов в КМ в них происходит синтез большого количества белковых фракций, входящих в состав мембран, синтез липидов и гема. Костномозговые ретикулоциты, каково бы ни было их абсолютное количество, образуют небольшой резерв красной крови. По ретикулоцитозу судят об эффективности эритропоэза. Содержание ретикулоцитов в норме 0,8-1,3, до 0,2-2 %.
Весь цикл развития от эритробласта до ретикулоцита составляет от 3-4 до 5-7 дней.
Собственно эритроцит. Эритроцит человека в норме имеет двояковогнутую, дискоидную форму. Плоский диск наиболее приспособлен к транспорту веществ из клетки и внутрь ее и к диффузии газов к центру клетки. Объем, соответствующий диску, имеет в 1,7 раза большую поверхность, чем такой же объем, соответствующий сфере и может умеренно изменяться без растяжения мембраны клетки.
Двояковогнутая форма эритроцита, эластичность, деформируемость мембраны позволяет клетке проходить через капилляры шириной 3 мкм, проникать в стенки синусоидов, возвращаясь к исходным параметрам (форме).
Эритроцит представлен клеточной мембраной, окружающей цитоплазму с включенными в нее гемоглобином и ферментами. Клеточная мембрана эритроцита представляет собой жидкий двойной липидный слой, в котором плавают белковые структуры. Взаимодействуя друг с другом белковые молекулы создают каркас мембраны, обеспечивающий ее прочность. Между белками и липидами существует прочная взаимосвязь. Липиды определяют подвижность белков и отвечают за пластичность и деформабельность.25-30 % мембранных белков составляет спектрин, существующий в виде димеров и тетрамеров. Спектрин, актин, протеины 4.1 и 4.9 образуют основу скелета мембраны, имеющего прочную жесткую структуру. Эритроцит теряет способность синтезировать белки и липиды после нескольких дней циркуляции в крови, поэтому особенно важно, чтобы на протяжении жизни эритроцита эти элементы были защищены.
С изменением цитоскелета эритроцита связаны некоторые формы гемолитических анемий - наследственные элиптоцитоз, пиропойкилоцитоз, отдельные варианты наследственного микросфероцитоза.
Время циркуляции эритроцита в кровотоке у взрослых составляет 100-120 дней.
По мере созревания эритроидных клеток в КМ происходит их гемоглобинизация.
Гемоглобин является дыхательным пигментом, относящимся к группе хромопротеидов. Он составляет около 95 % всей твердой части эритроцита. Гемоглобин это сложный белок, состоящий из белковой части - глобина и простетической части - гема. Молекула гемоглобина содержит 1 глобин и 4 гема. В каждом эритроците содержится около 289 млн. молекул гемоглобина. Гем представлен комплексом протопорфирина IX с железом. Одним из этапов синтеза гема является образование протопорфирина. Порфирины синтезируются во всех клетках организма, но основная масса в ядерных клетках эритроидного ростка КМ. Для синтеза одной молекулы гема необходимо 8 молекул аминокислоты глицина. В соединение с ним вступает янтарная кислота. Для осуществления этой связи в качестве кофермента необходим пиродоксаль-фосфат - производное витамина В6. В результате реакции образуется α-амино-β-кетоадипиновая кислота, которая очень нестойкая и при отщеплении от нее группы СО2 образуется δ-аминолевулиновая кислота (АЛК), из 2-х молекул которой образуется порфибилиноген (ПБГ). ПБГ под действием ферментов превращается в протопорфириногн III, а затем в протопорфириноген IX и протопорфирин IX. Протопорфирин IX при участии гемсинтетазы соединяется с Fe2+ и образуется гем. Протопорфирин состоит из 4-х пирроловых колец, в центре которых находится железо, соединенное с 4-мя атомами азота пирроловых колец двумя главными и двумя дополнительными связями. Так как координационное число железа равно 6, то остаются 2 неиспользованные координационные связи: с одной из них связывается глобин, а к другой присоединяется кислород или другие лиганды. Гем также синтезируется в мышцах для образования миоглобина, в клетках печени, для образования пероксидазы, цитохромов, каталазы.
Гем одинаков для всех видов гемоглобина, и различия в свойствах гемоглобина связаны с различием глобина.
Молекула гемоглобина представляет собой тетрамер, состоящий из 4-х полипептидных цепей. У человека продуцируются несколько типов цепей гемоглобина, которые при различных сочетаниях между собой определяют тип гемоглобина. У лиц с нормальным геномом встречается 4 типа полипептидных цепей глобина: α, β, γ и δ, отличающихся по составу и количеству аминокислот. В различные периоды развития человека отмечаются различные виды гемоглобина. В период онтогенеза выявляется гемоглобин Р (примитивный), к 8-ой неделе гестации 90 % составляет HbF. Он остается доминирующим весь период внутриутробного развития. Синтез HbA начинается с 9-ой недели гестации. К моменту рождения его содержание колеблется от 5 до 45 %.Затем его синтез резко увеличивается, а синтез HbF падает и к 6-ти месяцам его доля составляет менее 3 %, а HbA - 95-98 %. У взрослого к нормальным типам гемоглобина относятся HbA (α2 β2 - основной гемоглобин взрослого, 95-98 %), HbF (α2γ2 - фетальный гемоглобин, 0,5-1 %), HbA2 (α2δ2 - минорный гемоглобин взрослого, 1,5-3,5 %).
Эритроцит является высокоспециализированной клеткой, основная задача которой состоит в транспорте кислорода из легких в ткани и двуокиси углерода обратно в легкие. Эта функция осуществляется гемоглобином эритроцитов. Каждая молекула гемоглобина может обратимо связывать и транспортировать не более 4-х молекул кислорода.