

66. Обмен гистидина, глутамина, аспарагина, пролина.
Общая схема метаболизма глутамата и глутамина:
Ц







 ТК
аКГК
аКетоглутарамат
ТК
аКГК
аКетоглутарамат







 а-амино
NH3
карбомаил
кетокислоты
а-амино
NH3
карбомаил
кетокислоты


 бутират
фосфат
бутират
фосфат
аминокислоты










 N-ацетил
N-ацетил


 глутамат Глутамат
Глутамин
транспорт N2
глутамат Глутамат
Глутамин
транспорт N2









 ГАМК цис
о-про
карбомаил
ГАМК цис
о-про
карбомаил
 ГОМК
фосфат
ГОМК
фосфат
гамма-глу-амино фенилацетил


 гамма-глу-цис гли
кислота глутамин
гамма-глу-цис гли
кислота глутамин

гамма-глу-цис-гли аминокислота






с
 интез
из ГИС ПРОЛИН
интез
из ГИС ПРОЛИН
Глутаминовая кислота – сильное антитоксическое и антиоксидантное средство. Является центральным компонентом аминокислотного обмена, так как обеспечивает интеграцию белкового обмена с энергетическим (углеводным и липидным). Это осуществляется благодаря реакциям прямого и непрямого дезаминирования:
 ЦТК
ЦТК
Щ





 УК
аминокислота аКГК
NH3
мочевина
УК
аминокислота аКГК
NH3
мочевина
А

 СП
кетокислоты ГЛУ
СП
кетокислоты ГЛУ
 кетоновые тела
кетоновые тела
 фумарат глюкоза
фумарат глюкоза
мочевина
Глутамин играет роль антитоксического фактора. Это связано с тем, что при поступлении в клетку большого количества патологического материала, изменяется соотношение НАД и НАД*Н2, начинается конкуренция между субстратами за обладание НАД, в то время как концентрация НАД*Н2, возникает угроза энергетического голода тканей и дыхательная цепь переключается на ФАД-зависимое окисление именно благодаря глутамину, которая идёт на синтез эндогенного сукцинил-КоА.
Г


 ЛУ
ГАМК янтарный полуальдегид
сукцинилКоА ФАД-зависимый
ЛУ
ГАМК янтарный полуальдегид
сукцинилКоА ФАД-зависимый
COOH вход ДЦ
|
H
 OOC
– CH2
– CH2
– CH – COOH CH2
OOC
– CH2
– CH2
– CH – COOH CH2
| - CO2 |
NH2 CH2
ГЛУ |
CH2 – NH2
ГАМК
 аКГК
ГЛУ
COOH
COOH COOH
аКГК
ГЛУ
COOH
COOH COOH
. | HSKoA
|
|
| HSKoA
|
|


 CH2
CH2
CH2
ФАД (ДЦ)
CH2
CH2
CH2
ФАД (ДЦ)
| | |
CH2 CH2 CH2
| | |
H – C = O O = C – SKoA COOH
Янтарный полуальдегид сукцинилКоА сукцинат
Глутаминовая кислота является одним из немногих соединений в дополнение к глюкозе, которые служат энергетическим материалом для мозга (глутамин составляет 25% пула аминокислот мозга), поэтому в тканях мозга высока активность глутаматдекарбоксилазы:
HOOC – CH2 HOOC COOH COOH
| | | |
 CH2
а-декарбоксили
(CH2)
аКГК
ГЛУ
CH2
CH2
CH2
а-декарбоксили
(CH2)
аКГК
ГЛУ
CH2
CH2




 | рование
| |
| ДЦ
| рование
| |
| ДЦ
CH – NH2 CH2 CH CH2
 | |
| |
НАД*Н2
| |
| |
НАД*Н2
COOH NH2 O = C – H COOH
Глутамин ГАМК янтарный полуальдегид
НАД
COOH
|
CH2
|
CH2
|
ГОМК CH2OH
ГАМК и ГОМК – медиаторы торможения. Они оказывают тормозной эффект на синаптическую передачу. В общих реакциях (декарбоксилирование глутамина и трансаминирование ГАМК принимает участие ПФ, поэтому судороги, наблюдаемые при гиповитаминозе В6, связаны со снижением образования ГАМК в глутаматкарбоксилазной реакции.
Антиоксидантная активность глутамина заключается в том, что она входит в состав глутатиона. Поэтому при отравлении тяжёлыми металлами принимают глутамин во внутрь. Но ввиду слабой усвояемости чистого глутамина, его используют в качестве лекарственного препарата в виде солей (калиевой соли глутамина). Глутамин имеет вкус куриного бульона.
Метаболизм аспарагина.
Синтез аспарагина осуществляется реакцией трансаминирования:
COOH COOH COOH COOH
| | | |
CH2 (CH2) АсАТ CH2 (CH2)

 |
+ |
| + |
|
+ |
| + |
C = O CH – NH2 CH – NH2 C = O
| | | |
COOH COOH COOH COOH
ЩУК глутамин аспарагин аКГК
Аспартат утилизируется:
-
В ГНГ (глюкогенная аминокислота)
-
Синтез мочевины
-
Синтез пуринов и пиримидинов
-
Образование в-аланина или а-аланина
COOH CH3 – CH – COOH

 |
|
COOH анзерин
|
|
COOH анзерин
CH2 NH2 |

 |
CH2
|
CH2
CH – NH2 - CO2 | карнозин
| CH2 – NH2
COOH в-аланин
В-аланин входит в состав пантогеновой кислоты, идущей на синтез HS-KoA. Опухолевые ткани не способны воспроизводить аспарагин, поэтому введение аспарагиназы приводит к торможению роста опухоли.
-
Трансаминирование аспарагина даёт ЩУК
 ЩУК
сукцинат, который наполняет эндогенный
пул янтарной кислоты.
ЩУК
сукцинат, который наполняет эндогенный
пул янтарной кислоты.
-
Синтез аспарагина. Нужно отметить, что глицин и аспарагин в тканях подвергаются сочетанному трансаминированию и дезаминированию:
O O HOOC – C = O
|| || R |

 R – C – COOH + HOOC – CH – CH2
– C | +
CH2
R – C – COOH + HOOC – CH – CH2
– C | +
CH2
Кетокислота | | CH – NH2 |
NH2 NH2 | C = O
Аспсрагин COOH |
Аминокислота NH2
а-кетосукцинамовая
кислота
NH2 COOH
| |
C = O CH2
| + HOH | + NH3
C H2
C
= O
H2
C
= O
| СО-оксидаза |
C = O COOH
| ЩУК
COOH
а-кетосукцинамовая кислота
То же самое возможно и с глицином, при этом образуется а-кетоглутарамовая кислота.
-
Аспартат является нейротрансмиттером ЦНС (в виде препарата натриевой соли аспартата применяется для лечения заболеваний сердца).
Метаболизм гистидина.
Известны 3 пути метаболизма гистидина:
1.







 N
N
N
N








 CH2
– CH – COOH
CH2
– CH2
– NH2
CH2
– CH – COOH
CH2
– CH2
– NH2
| - CO2
NH NH2 NH гистамин
O




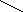






 N
+
HOH N
//
N
+
HOH N
//




 CH2
– CH2
– NH2
NH3
+ CH2
– C + H2O2
CH2
– CH2
– NH2
NH3
+ CH2
– C + H2O2
NH диаминооксидаза NH \
альдегид гистидина H
2



 .
.
N



 CH2
– CH – COOH + H2N
– CH2
– CH2
– COOH
CH2
– CH – COOH + H2N
– CH2
– CH2
– COOH
NH | в-аланин - НОН





 гистидин
NH2
гистидин
NH2


 N
N
N
N



 CH2
– CH – COOH
CH2
– CH – COOH
CH2
– CH – COOH
CH2
– CH – COOH




 |
|
|
|
NH NH N NH
| | |
H2N – CH2 – CH2 – C = O CH3 CO
карнозин |
анзерин (CH2)2 – NH2 Внутреннее дезаминирование:
3. 










 N
гистидинаммиаклиаза
N
N
гистидинаммиаклиаза
N





 CH2
– CH – COOH
CH
= CH – COOH
CH2
– CH – COOH
CH
= CH – COOH
NH | - NH3 NH
NH2


 имидазолилпропионат
ГЛУ
имидазолилпропионат
ГЛУ
Метаболизм пролина.
Пролин синтезируется из глутамата:
O
НАДФ*Н2 НАДФ //
H

 OOC
– CH2
– CH2
– CH – COOH HOOC – CH2
– (CH2)
– C
OOC
– CH2
– CH2
– CH – COOH HOOC – CH2
– (CH2)
– C
- НОН гамма-полуальдегид \





 H
H
г

 лутаматдегидрогеназа
лутаматдегидрогеназа


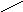

 COOH
O = COOH
COOH
O = COOH




 - НОН
NH
пролиноксидаза
NH
- НОН
NH
пролиноксидаза
NH
пролин 5-оксопролин
оксипролиназа COOH COOH


 O = COOH
| | GSH
O = COOH
| | GSH
 NH
CH2
– CH2
– CH – NH2
NH
CH2
– CH2
– CH – NH2
5-оксопролин АТФ АДФ+Фн глутамат