

7. Плазмиды.
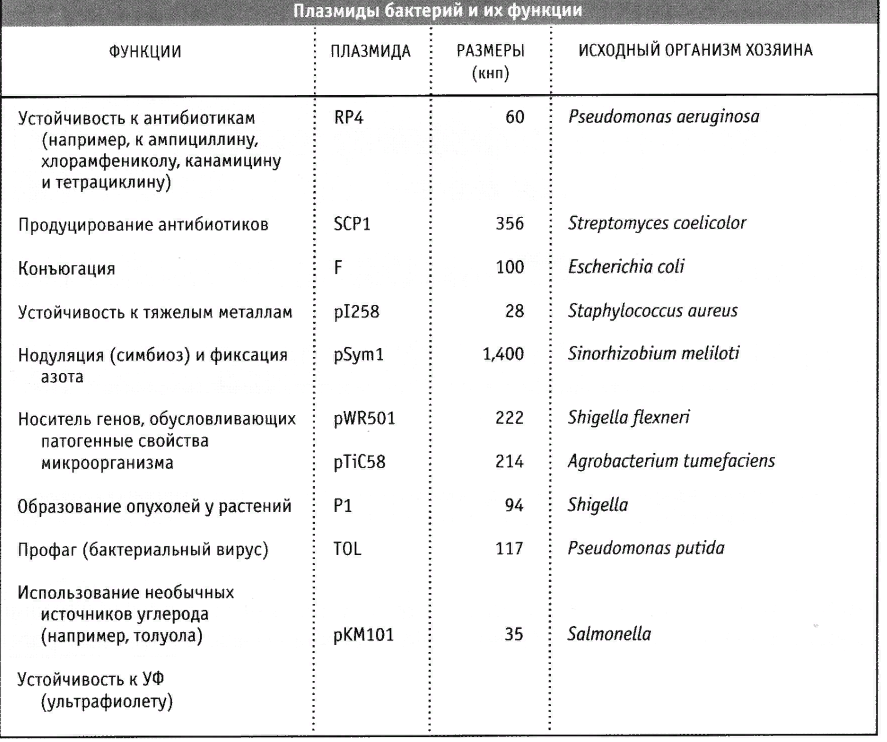
Они могут передаваться от одной клетки к другой путем горизонтального переноса и обеспечивают некоторые преимущества для клеток-хозяев.
Плазмиды содержат важную область генов или локусов, участвующих в их репликации и контроле. Кроме того, плазмиды могут содержать гены, которые можно назвать «необязательными», однако они могут играть важную роль в жизни клетки-хозяина.
Итак, плазмиды содержат:
-
точку ori (последовательность нуклеотидов, с которой начинается репликация);
-
ген, кодирующий Rep-белок, участвующий в инициации репликации плазмиды;
-
гены, кодирующие регуляторные белки репликации.
Нередко они могут передаваться от одной бактерии к другой того, таким образом, служат средством горизонтального переноса генов. Перенос плазмиды в клетку может осуществляться двумя путями: либо при непосредственном контакте клетки-хозяина с другой клеткой в процессе конъюгации, либо путём трансформации, то есть захвата экзогенной ДНК из внешней среды.
8. Клеточная стенка грамположительных бактерий.
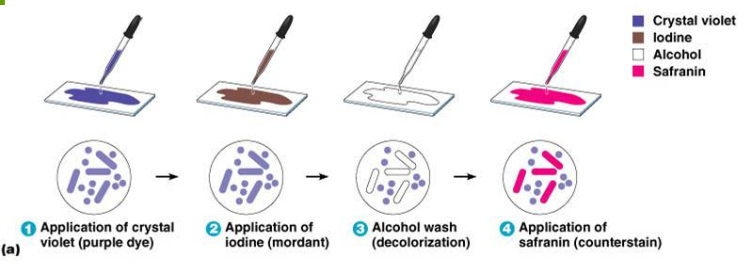
Суть метода Грама в том, что исследуемую среду обрабатывают анилиновым красителем, фиксируют йодом, а затем смывают спиртом. При этом грамотрицательные бактерии обесцвечиваются, а грамположительные приобретают синюю окраску.
Это происходит из-за наличия внешней мембраны, препятствующей проникновению красителя внутрь клетки.
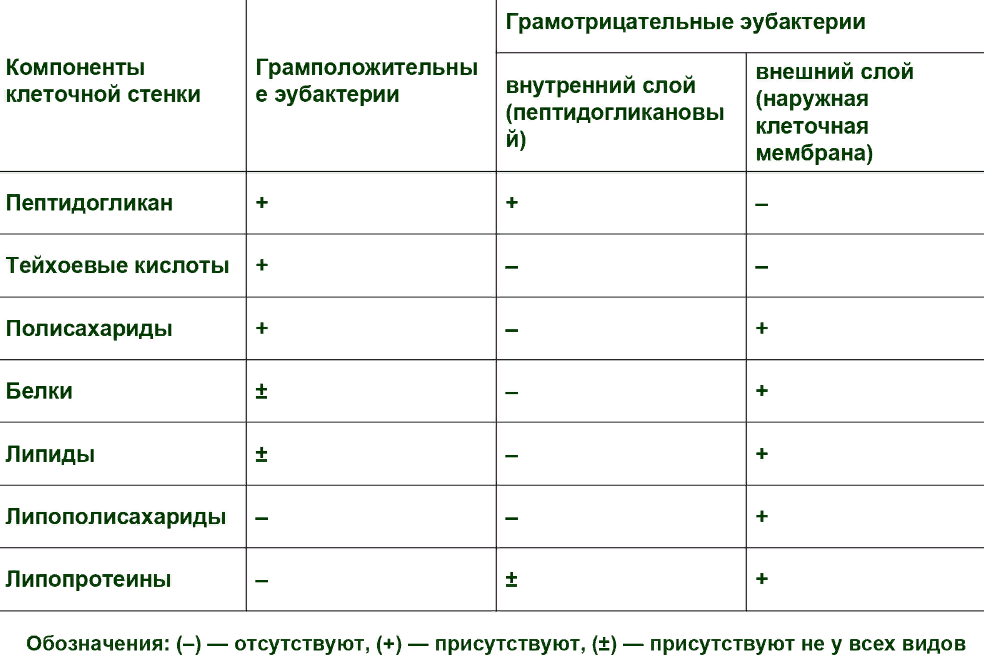
Клеточная стенка грамположительных бактерий плотно прилегает к цитоплазматической мембране.
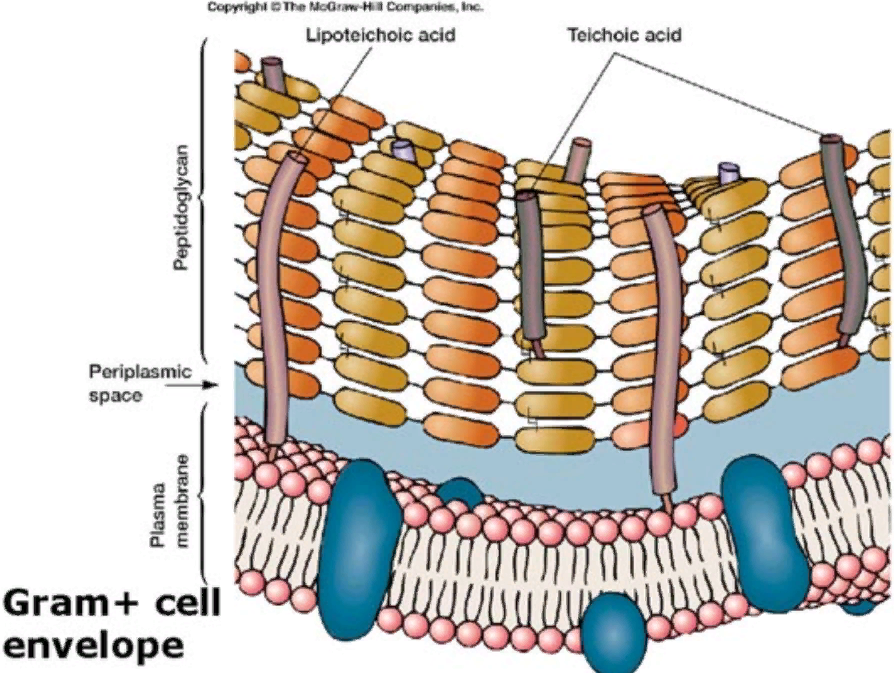
Опорный скелет бактериальной стенки состоит из пептидогликана или муреин. Тетрапептидные боковые цепи мурамовой кислоты связаны между собой межпептидными (например, пентаглициновыми) цепочками. Участвующие в образовании таких структур аминокислоты варьируют от вида к виду. Содержание белков невелико. Характерная особенность - наличие тейхоевых кислот; это цепи, состоящие из 8-50 остатков глицерола или рибитола, связанных между собой фосфатными мостиками. Некоторые из тейхоевых кислот содержат эритритол или маннитол. Тейхоевые кислоты, вероятно, через фосфат связаны с муреином по типу амида.

Эта макромолекула - гетерополимер, построенный из цепочек, в которых чередуются остатки N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенные между собой гликозидными связями. Такие неразветвленные гетерополимерные цепи образуют основу муреина. Остатки мурамомвой кислоты через лактильные группы соединены пептидной связью с аминокислотами. К типичным аминокислотам муреина относятся L-аланин, D-глутаминовая кислота, мезо-диаминопимелиновая кислота или L-лизин и D-аланин.

Диаминокислоты мезо- (или LL-) диаминопимелиновая кислота и L-лизин играют большую роль в межмолекулярных сшивках, так как образуют пептидные связи с участием обеих аминогрупп и, таким образом, могут связать две гетерополимерные цепи между собой (рис. 2). Место диаминопимелиновой кислоты или лизина могут занять орнитин или диаминомасляная кислота. Пептидными мостиками гетерополимерные цепи связаны между собой в мешкообразную гигантскую молекулу - муреиновый мешок.
Особого внимания заслуживает то, что в бактериальной стенке содержатся структуры и вещества, которых нет у животных и растений: таковы, например, чередующаяся последовательность N-ацетилглюкозамина и N-ацетилмурамовой кислоты; не встречающаяся в составе белков мезо-диаминопимелиновая кислота; D-формы аланина и глутаминовой кислоты. Эти структурные элементы составляют ахиллесову пяту бактерий, используемую врачами в борьбе с инфекцией. По компонентам и структуре клеточной стенки и биохимическим механизмам ее синтеза бактерии коренным образом отличаются от животных и растений. Поэтому лекарственные препараты, специфически воздействующие только на бактериальные стенки и на процесс их синтеза, должны быть безвредными для высших организмов.