

5. Регуляция цикла трикарбоновых кислот
Едва ли можно сомневаться в том, что в большинстве тканей, в которых основная функция цикла лимонной кислоты - обеспечение энергией, дыхательный контроль, осуществляемый при функционировании дыхательной цепи и окислительного фосфорилирования, является определяющим фактором при регуляции активности рассматриваемого цикла. Активность этого цикла непосредственно связана с поступлением окисленных кофакторов дегидрогеназ (например, НAД), которое в свою очередь зависит от доступности AДФ и, в конечном счете, от скорости потребления АТФ. Свойства ряда ферментов этого цикла указывают на то, что кроме обшей регуляции существует также регуляция на уровне самого цикла. В клетках головного мозга, в которых ацетил-КоА образуется в основном из углеводов, регуляция цикла лимонной кислоты может происходить на стадии, катализируемой пируватдегидрогеназой. В самом цикле регуляция может осуществляться путем аллостерического ингибирования цитратсинтетазы при действии АТФ или ацил-КоА-производных длинноцепочечных жирных кислот. Митохондриальная НAД-зависимая изоцитратдегидрогеназа аллостерически активируется AДФ и ингибируется АТФ и НAДH2.
а-Кетоглутаратдегидрогеназный комплекс регулируется, по-видимому, аналогично пируватдегидрогеназе. Сукцинатдегидрогеназа ингибируется оксалоацетатом, а образование оксалоацетата в малатдегидрогеназной реакции зависит от соотношения [НAДH2]/[НAД+]. Поскольку величина Км цитратсинтазы для оксалоацетата такого же порядка, что и величина внутримитохондриальной концентрации оксалоацетата, концентрация последнего, по видимому, играет определенную роль в регуляции скорости образования цитрата. Какие из вышеперечисленных механизмов регуляции в самом цикле функционируют in vivo, пока еще не ясно [8].
6.Пентозофосфатный цикл (гексозомонофосфатный путь метаболизма глюкозы).
Открытие пути прямого окисления углеводов, или, как его называют пентозофосфатного цикла, принадлежит О. Варбургу, Ф. Липману
Ф. Дикенсу и В. А. Энгельгарду [1]. Расхождение путей окисления углеводов -классического (цикл трикарбоновых кислот, или цикл Кребса) и пентозофосфатного - начинается со стадии образования гексозомонофосфата. Если глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат, который фосфорилируется второй раз и превращается во фруктозо-1,6-бисфосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути.
Если второго фосфорилирования гексозо-6-монофосфата не происходит, то фосфорилированная глюкоза может подвергаться прямому окислению до фосфопентоз (рибозо-5-фосфат и ксилулозо-5-фосфат). В норме доля пентозофосфатного пути в количественном превращении глюкозы обычно невелика, варьирует у разных организмов и зависит от типа ткани и ее функционального состояния.
Пентозофосфатный цикл протекает в цитоплазме и состоит из 2-х этапов:
1. Окислительный.
2.Неокислительный.
Значение окислительного этапа пентозофосфатного цикла.
1.Происходит прямое окисление глюкозофосфата без участия кислорода.
2.Этот этап является одним из главных источников НАДФН2для клетки. Образуется этот НАДФН2в цитоплазме, поэтому он не передает свой водород по системе митохондриального окисления на кислород и АТФ не образуется. Он отдает свой водород на синтез жирных кислот, холестерина и других стероидов, а также на монооксигеназные реакции.
3.На 1-м этапепентозофосфатного цикла образуется СО2- один из конечных продуктов метаболизма без участия кислорода.
4.Образуются пентозы. Эти пентозы являются строительным материалом для синтеза нуклеотидов, коферментов и некоторых других веществ.
Значение неокислительного пути.
1.Обеспечивает завершение 1-го этапа (утилизирует продукты 1-го этапа).
2.Является источником моносахаридов с разным числом углеродных атомов. Это строительный материал для разных синтезов, в том числе для синтезов различных олигосахаридов, которые входят в состав различных клеточных рецепторов.
3.Образующийся ФГА является точкой сопряжения между пентозофосфатным циклом и некоторыми другими путями метаболизма. Например: ФГА может восстанавливаться до фосфоглицерина, который нужен для синтеза жиров. Фосфоглицерин может окисляться до ФГА. ФГА также образуется в процессе гликолиза и является общим метаболитом.
РЕГУЛЯЦИЯ ПЕНТОЗОФОСФАТНОГО ПУТИ
Лимитирующим звеном пентозофосфатного путиокисления углеводов является реакция окисления 6-фосфоглюконовой кислоты, катализируемая
6-фосфоглюконатдегидрогеназой.
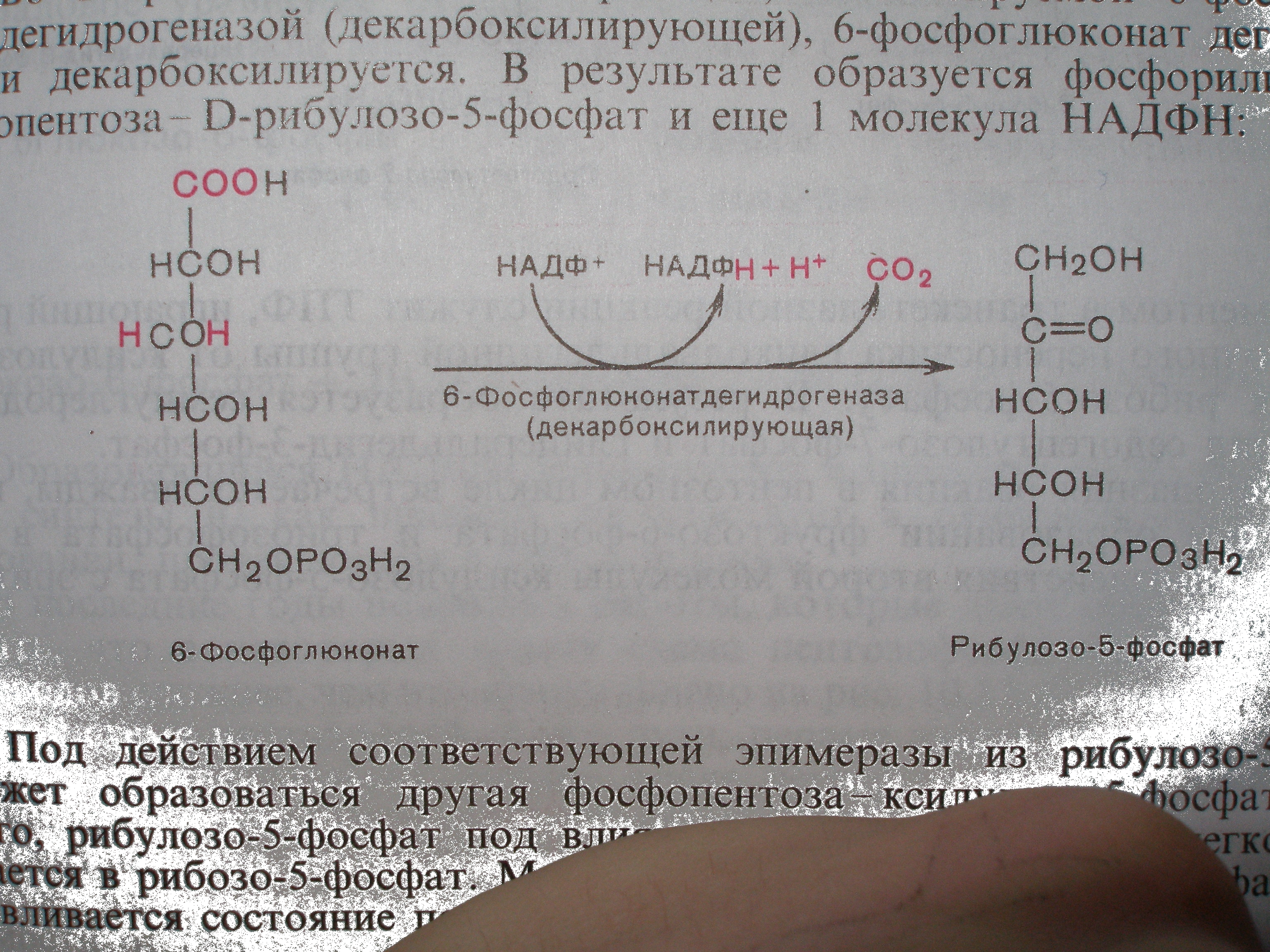
Vmaxэтого фермента меньше, чем Vmaxлюбого другого ферментапентозофосфатного цикла. Поэтому он определяет собой Vmaxвсегопентозофосфатного циклав целом и является лимитирующим ферментом. Но он не является ключевым, потому что не является регуляторным.
Регуляторный фермент пентозофосфатного цикла - это глюкозо-6-
фосфатдегидрогеназа.
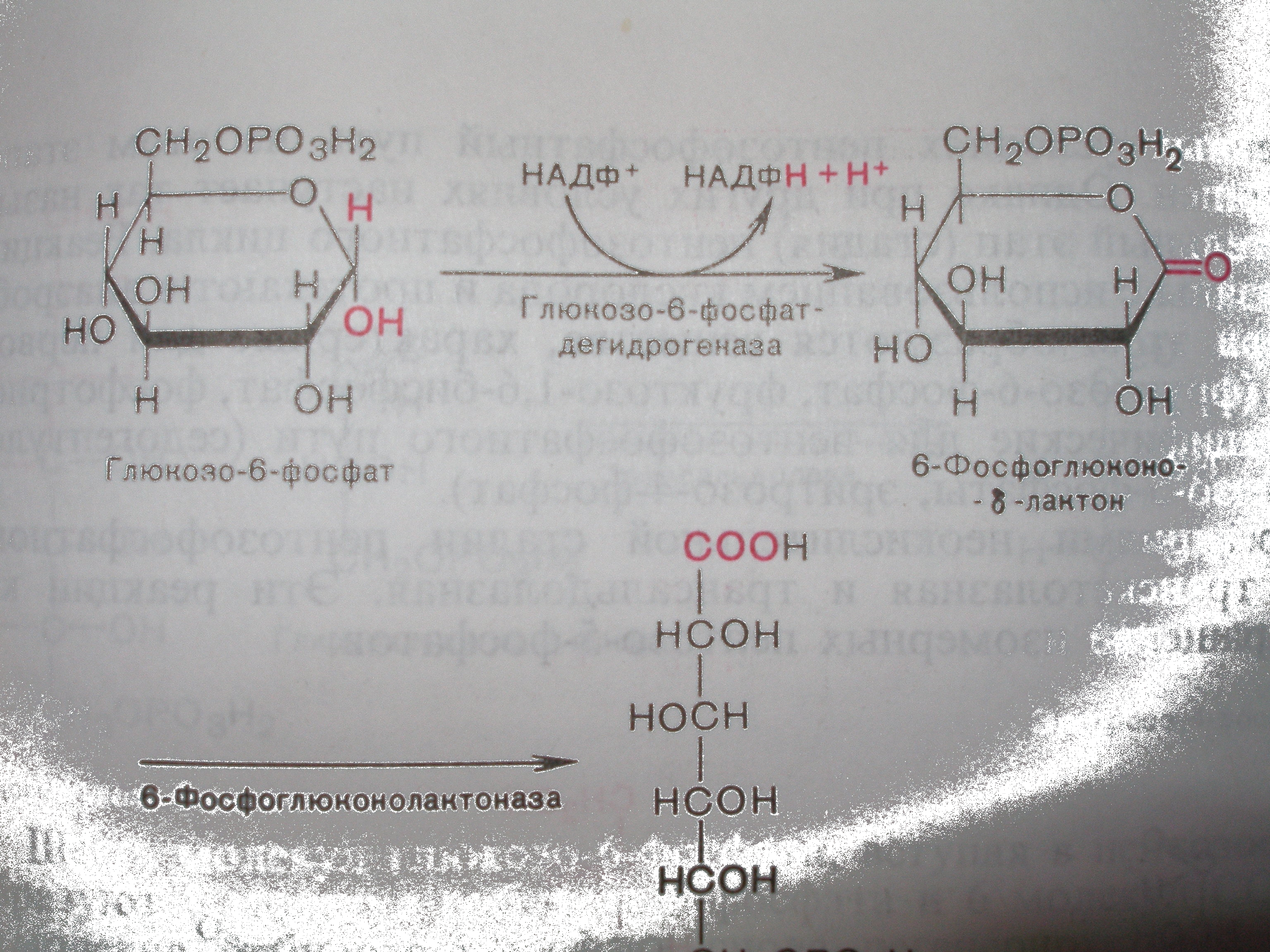
Его Vmaxбольше, чем у 6-фосфоглюконатдегидрогеназы, поэтому он не является лимитирующим. Но он ингибируется избытком АТФ и своего продукта НАДН2. Значит, в определенных условиях его Vmaxможет стать меньше, чем Vmax6-фосфоглюконатдегидрогеназы. В этих условиях лимитирующая роль переходит к глюкозо-6-фосфатдегидрогеназе.
Таким образом, роль ключевого фермента пентозофосфатного циклавыполняют сразу 2 фермента - обе дегидрогеназы, которые разделяют между собой функции ключевого и лимитирующего ферментов. Эти ключевые ферменты являются "пунктами вторичного контроля"пентозофосфатного цикла. Скоростьпентозофосфатного циклаопределяется либо 6-фосфоглюконатдегидрогеназой, либо (когда много НАДФН2 и АТФ) - глюкозо-6-фосфатдегидрогеназой. Реакциипентозофосфатного цикларегулируются генетически на уровне биосинтеза этих ферментов.
Пентозофосфатный циклсуществует не во всех типах клеток. Наиболее интенсивнопентозофосфатный циклпротекает в печени, эритроцитах, надпочечниках, половых железах, жировой ткани и молочной железе. Но даже в этих тканях ГМФ-путем расщепляется не более 25-30% глюкозы. В этих тканяхпентозофосфатный циклимеет такое значение, потому что клеткам нужно много НАДФН2. Например: В жировой ткани с большой скоростью идет синтез жиров. В печени - синтез жиров, холестерина. В коре надпочечников и в половых клетках синтезируются стероидные гормоны. Для этого нужно много НАДФН2 [2,6,8].
Регуляция на генетическом уровнеидет медленно, требуется время для проявления эффекта этого типа регуляции. Автономная регуляция приводит к практически мгновенному эффекту[6].
Пентозофосфатный цикл в эритроцитах нужен для обеспечения НАДФН2, но не для синтезов, а для защиты от токсических эффектов кислорода. В эритроцитах многие ферменты имеют в активных центрах
SH-группы, которые могут окисляться под действием кислорода. Накопленный при распаде глюкозы в пентозофосфатном циклеНАДФН2позволяет регенерировать эти SH-группы.