

ВОЕННО-МЕДИЦИНСКАЯ АКАДЕМИЯ
Экз №__
Кафедра клинической биохимии и лабораторной диагностики
«УТВЕРЖДАЮ»
ИО начальника кафедры
клинической биохимии и
лабораторной диагностики
полковник медицинской службы
В.ПАСТУШЕНКОВ
«___» _____________ 2008 г.
доцент кафедры клинической биохимии и лабораторной диагностики
кандидат биологических наук доцент Е.ШЕЛЕПИНА
_____________________________________________________________________
должность, ученая степень, ученое звание, воинское звание, инициал имени, фамилия автора (авторов)
ЛЕКЦИЯ № 12
_________________________________________
(номер по тематическому плану изучения дисциплины)
по дисциплине: «Биохимия»
___________________________________________________________
(наименование учебной дисциплины)
на тему: «БИОСИНТЕЗ ЛИПИДОВ»
________________________________________________
(наименование темы занятий по тематическому плану изучения дисциплины)
с курсантами и студентами 2 курса факультетов подготовки врачей
(военно-медицинских специалистов иностранных армий)
Обсуждена и одобрена на заседании кафедры
«____» ____________ 200___ г.
Протокол №______
Уточнено (дополнено):
«____» ____________ 200___ г.
_____________________________________
(воинское звание, подпись, инициал имени, фамилия)
Содержание.
Ведение…………………………………………………….…………………………………стр.2
Синтез жирных кислот в организме………………………………………………………. стр.2
Образование ненасыщенных жирных кислот……………………………………………..стр.5
Простагландины……………………………………………………………………………. стр.6
Биосинтез триглицеридов…………………………………………………………………..стр.7
Биосинтез глицерофосфолипидов (фосфоглицеридов)…………………………………...стр.9
Биосинтез холестерина…………………………………………………………………….стр.11
Обмен этанола……………………………………………………...………………………стр.14
Заключение……………………………………………………...………………………….стр.14
-1-
Введение.
Способность животных и человека запасать углеводы в форме гликогена (в печени и мышцах) довольно ограничена. Так, в организме может содержаться лишь несколько сотен граммов гликогена, которых хватит для обеспечения потребности организма в энергии не более чем на 12 часов. На долю же триацилглицеридов в организме человека весом 70 кг приходится около 12 кг. Накопленной в них энергии достаточно для поддержания основного обмена человека в течение 8 недель. Когда углеводы потребляются в избытке и организм уже не способен откладывать их в форме гликогена, они превращаются в триацилглицериды, которые в больших количествах могут накапливаться в клетках жировой ткани. Больше всего таких клеток находится в подкожно жировой клетчатке и в брюшной полости.
Полярные липиды (фосфолипиды и сфинголипиды) в организме животных не запасаются, но они также постоянно синтезируются для восполнения потерь, обусловленных разрушением мембран в ходе метаболических превращений. Например, время полужизни молекул мембранных фосфолипидов в печени крысы составляет менее 3 суток.
Важным процессом является обмен и биосинтез холестерина и других стероидов в организме. Известно, что из холестерина в организме образуются многочисленные биологически активные стероиды: половые гормоны, кортикостероиды, желчные кислоты, провитамин Д3 и др. холестерин является составной частью биологических мембран.
1.Синтез жирных кислот в организме.
В настоящее время в результате работ Уокила, Линена, Брэди и других ученых в достаточной степени изучен механизм биосинтеза жирных кислот в организме животных и человека, а также катализирующие этот процесс системы. В митохондриях же в основном происходит удлинение существующих цепей жирных кислот. Например, установлено, что в цитоплазме печёночных клеток синтезируется главным образом пальмитиновая кислота, а в митохондриях печёночных клеток на основе уже синтезированной в цитоплазме клетки пальмитиновой кислоты или на основе жирных кислот экзогенного происхождения, т.е. поступивших из кишечника, образуются жирные кислоты, содержащие 18, 20 и 22 углеродных атома. При этом реакции синтеза жирных кислот в митохондриях по существу являются обратными реакциями окисления жирных кислот.
Внемитохондриальный же синтез (основной, главный) жирных кислот по своему механизму резко отличается от процесса их окисления.
Строительным блоком для синтеза жирных кислот в цитоплазме клетки служит ацетил-КоА, который в основном ведёт своё происхождение от митохондриального ацетил-КоА. Установлено также, что для синтеза жирных кислот важно наличие в цитоплазме двуокиси углерода или иона бикарбоната. Кроме того, было выявлено, что цитрат стимулирует синтез жирных кислот в цитоплазме клетки. Известно, что образующийся в митохондриях в процессе окислительного декарбоксилирования ацетил-КоА не может диффундировать в цитоплазму клетки, ибо митохондриальная мембрана не проницаема для данного субстрата. Показано, что митохондриальный ацетил-КоА взаимодействует с оксалоацетатом, в результате образуется цитрат, который свободно проникает в цитоплазму клетки, где расщепляется до ацетил-КоА и оксалоацетата (слайд 2):
в митохондриях
COOH
CH3-CO-S-KoA +HOOC-CO-CH2-COOH --→ HOOC-CH2-C- CH2-COOH + HS-KoA
+H2O
ацетил-КоА оксалоацетат OH
лимонная кислота
-2-
в цитоплазме
COOH
HOOC-CH2-C- CH2-COOH + АТФ+ HS-KoA --→ CH3-CO-S-KoA +
OH ацетил-КоА
лимонная кислота
+ HOOC-CO-CH2-COOH + АДФ +Н3РО4
оксалоацетат
Следовательно, в данном случае цитрат выступает в роли переносчика ацетильного радикала.
Есть ещё один путь переноса внутримитохондриального ацетил-КоА в цитоплазму клетки. Это – путь с участием карнитина. Выше указывалось, что карнитин играет роль переносчика ацильных групп из цитоплазмы и митохондрий при окислении жирных кислот. По-видимому, он может выполнять эту роль и в обратном процессе, т. е. в переносе ацильных радикалов, в том числе ацильного радикала из митохондрий в цитоплазму клетки. Однако, когда речь идёт о синтезе жирных кислот, данный путь переноса ацетил-КоА не является главным.
Важнейшим шагом в понимании процесса синтеза жирных кислот было открытие фермента ацетил-КоА-карбоксилазы. Этот сложный фермент, содержащий биотин, катализирует АТФ-зависимый синтез малонил-КоА (НООС-СН2-СО ~S-КоА) из ацетил-КоА и СО2 (слайд 3).
Данная реакция протекает в два этапа:
СО + АТФ + биотин-энзим ----→ карбоксибиотин-энзим + АДФ + Н3РО4 (1)
Карбоксибиотин-энзим + CH3-CO-S-KoA ----→
Ацетил-КоА
----→ НООС-СН-СО-S-КоА + биотон-энзим (2)
Малонил-КоА
Установлено, что функцию активатора ацетил-КоА-карбоксилазной реакции выполняет цитрат.
Таким образом, малонил-КоА представляет собой первый специфический продукт биосинтеза жирных кислот. В присутствии соответствующей ферментативной системы малонил-КоА (который в свою очередь, образуется из ацетил-КоА) быстро превращается в жирные кислоты.
Ферментная система, синтезирующая высшие жирные кислоты, состоит из нескольких ферментов, определённым образом связанных между собой.
В настоящее время процесс синтеза жирных кислот детально изучен Е. cоli и некоторых других микроорганизмов. Мультиферментный комплекс, именуемый синтетазой жирных кислот (слайд 4), состоит у Е. соli из семи ферментов, связаных с так называемым ацилпереносящим белком (АПБ) (слайд 5). Этот белок относительно термостабилен, имеет свободную HS-группу и вовлекается в процесс синтеза высших жирных кислот практически на всех его этапах (слайд 6). Относительная молекулярная масса АПБ составляет около 10000 дальтон.
Ниже приводится последовательность происходящих при синтезе жирных кислот реакций:
Условно мультиэнзимный комплекс синтазы жирных кислот (Е) представлен следующим образом:
 Стекло
№1, HS-Цис
Стекло
№1, HS-Цис
 №2 Е
№2 Е
HS-Фп-АПБ
где Фп – фосфопантотеновая кислота, являющаяся простетической группой АПБ. Он необходим для связывания малонил-КоА
-3-
1
 этап HS-Цис
СН3
ацетил-КоА-АПБ
трансфераза СН3
-СО S-Цис
этап HS-Цис
СН3
ацетил-КоА-АПБ
трансфераза СН3
-СО S-Цис
 Е +
-----------------------------------→
Е
Е +
-----------------------------------→
Е
HS-Фп-АПБ С=О -HSKoA HS-Фп-АПБ
SКоА
Ацетил-КоА
2 этап COOH
 СН3 -СО S-Цис
малонил-КоА-АПБ
трансфераза
СН3 -СО S-Цис
малонил-КоА-АПБ
трансфераза
 Е + СН2
-----------------------------------→
Е + СН2
-----------------------------------→
HS-Фп-АПБ
С=О
SКоА
малонил-КоА
 СН3
-СО
S-Цис
СН3
-СО
S-Цис
 -------→
Е + HS-KoA
-------→
Е + HS-KoA
СОО-СН2СО S-Фп-АПБ

 АцетилS-Цис
3-кетоацил-АПБ-синтаза
HS-Цис
АцетилS-Цис
3-кетоацил-АПБ-синтаза
HS-Цис
 Е
-----------------------------------→
Е
Е
-----------------------------------→
Е
М алонилS-Фп-АПБ
СН3-СО-СН2-СОS-Фп-АПБ
алонилS-Фп-АПБ
СН3-СО-СН2-СОS-Фп-АПБ
ацетоацетилS-Фп-Е
3 этап (восстановление)
 СН HS-Цис
СН HS-Цис

 HS-Цис
3-кетоацил-АПБ-редуктаза
HS-Цис
3-кетоацил-АПБ-редуктаза

 Е
----------------------------------→СН-ОН
Е
Е
----------------------------------→СН-ОН
Е
С Н3-СО-СН2-СОS-Фп-АПБ
НАДФН2
НАДФ
Н3-СО-СН2-СОS-Фп-АПБ
НАДФН2
НАДФ
СН2-СОS-Фп-АПБ
4 этап (дегидратация):
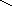 HS-Цис
β-гидроксибутирил-АПБ-дегидратаза
HS-Цис
β-гидроксибутирил-АПБ-дегидратаза
 Е
------------------------------------------------→
Е
------------------------------------------------→
СН3-СН-СН2-СОS-Фп-АПБ
׀
ОН
 HS-Цис
HS-Цис
-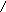 ---→
Е
---→
Е
СН3-СН=СН-СОS-Фп-АПБ
Кротонил-АПБ
5 этап
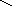
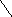 HS-Цис
енол-АПБ-редуктаза СН3
HS-Цис
HS-Цис
енол-АПБ-редуктаза СН3
HS-Цис
 Е
-------------------------→
׀
Е
-------------------------→
׀
С Н3-СН=СН-СОS-Фп-АПБ
НАДФН2
НАДФ
СН2
Е
Н3-СН=СН-СОS-Фп-АПБ
НАДФН2
НАДФ
СН2
Е
׀
СН2-СОS-Фп-АПБ
6 этап (перенос):

 HS-Цис ТФ
Бутирил
S-Цис
HS-Цис ТФ
Бутирил
S-Цис

 Е ------→
Е
Е ------→
Е
Бутирил S-Фп-АПБ НS-Фп-АПБ
7 этап новый цикл реакций (наращивание цепи жирной кислоты)
СООН
Б
 утирил
S-Цис ׀
малонил-КоА-АПБ-трансфераза
Бутирил
S-Цис
утирил
S-Цис ׀
малонил-КоА-АПБ-трансфераза
Бутирил
S-Цис

 Е + СН2
--------------------------------------→
Е
Е + СН2
--------------------------------------→
Е
НS-Фп-АПБ ׀ СООН- СН2-СОS-Фп-АПБ
СОS-КоА
Малонил-КоА
-4-

 Бутирил
S-Цис -СО2
НS-Цис
Бутирил
S-Цис -СО2
НS-Цис

 Е ----→
Е
Е ----→
Е
СООН- СН2-СОS-Фп-АПБ СН3-(СН2)2-СО-СН2-СОS-Фп-АПБ


 Деацилаза
Н2О
Деацилаза
Н2О
Повтор циклов ПальмитилS-АПБ-Е
По сравнению с -окислением биосинтез жирных кислот имеет ряд характерных особенностей: синтез жирных кислот в основном осуществляется в цитоплазме клетки, а окисление – в митохондриях; участие в процессе биосинтеза жирных кислот малонил-КоА, который образуется путём связывания СО2 (в присутствии биотин-фермента и АТФ) с ацетил-КоА; на всех этапах синтеза жирных кислот принимает участие ацилпереносящий белок (HS-АПБ). Установлена необходимость для синтеза жирных кислот кофермента НАДФ-Н2. Последний в организме образуется частью (на 50 процентов) в реакциях пентозного цикла (гексозомонофосфатного «шунта»), частью – в результате восстановления НАДФ малатом (яблочная кислота + НАДФ ----→
пировиноградная кислота + СО2 + НАДФ-Н2); восстановление двойной связи в еноил-АПБ-редуктазной реакции происходит при участии НАДФ-Н2 и фермента, простетической группой которого является флавинмононуклеотид (ФМН); в процессе синтеза жирных кислот образуются гидроксипроизводные, относящиеся по своей конфигурации к Д-ряду жирных кислот, а при окислении жирных кислот – гидроксипроизводные L-ряда.
В тканях животных не удалось обнаружить АПБ. В выделенном из печени мультиферментном комплексе, катализирующем синтез жирных кислот, имеются две свободные SH-группы, одна из которых, как и АПБ, принадлежит фосфорилированному пантетеину, другая цистеину. Считают, что пантетеиновая SH-группа комплекса выполняет такую же роль, как и SH-группа АПБ, т. е. Участвует в связывании и переносе ацильного радикала.