

11. Обмен липидов
Роль липидов в питании
Липиды являются обязательной составной частью сбалансированного пищевого рациона человека. Принято считать, что при сбалансированном питании соотношение белков, липидов и углеводов в пищевом рационе составляет примерно 1:1:4. В среднем в организм взрослого человека с пищей ежесуточно поступает около 80г жиров животного и растительного происхождения (А.А.Покровский и др.1978). В пожилом возрасте, а также при малой физической нагрузке потребность в жирах снижается. В условиях холодного климата и при тяжелой физической работе потребность в жирах у человека повышается. Значение жиров как пищевого продукта весьма многообразно. Прежде всего жиры в питании человека имеют важное энергетическое значение. Высокая калорийность жиров по сравнению с белками и углеводами придает им особую пищевую ценность при расходовании организмом больших количеств энергии. Известно, что 1г жиров при окислении в организме дает 9.3 ккал, тогда как 1г белка или углеводов дает 4.1 ккал. Следует также помнить, что жиры являются растворителями витаминов А, Д, Е, и др., в связи с чем обеспеченность организма этими витаминами в значительной степени зависит от поступления жиров в составе пищи. Кроме того, с жирами в организм вводятся некоторые полиненасыщенные кислоты (линолевая, линоленовая, арахидоновая кислоты), которые относят к категории незаменимых жирных кислот, ибо ткани человека и ряда животных потеряли способность синтезировать их. Эти кислоты условно объединены в группу под названием витамин «F».
Наконец, с жирами организм получает комплекс биологически активных веществ таких, как фосфолипиды, стерины и др., играющих важную роль в липидном обмене.
III. Переваривание и всасывание лпипидов (переваривание липидов-слайд №7)
Расщепление жиров в желудочно-кишечном тракте. Слюна не содержит расщепляющих жиры ферментов. Следовательно, в полости рта жиры не подвергаются никаким изменениям. У взрослых людей жиры проходят через желудок также без особых изменений, поскольку содержащаяся в небольшом количестве в желудочном соке взрослого человека и млекопитающих липаза малоактивна. Величина pH желудочного сока около 1.5, а оптимальное значение pH для желудочной липазы находится в пределах 5.5 – 7.5 Кроме того, липаза может активно гидролизировать только предварительно эмульгированные жиры, в желудке же отсутствуют условия для эмульгирования жиров. Переваривание жиров в полости желудка играет важную роль в процессе пищеварения у детей, особенно у детей грудного возраста. Известно, что pH желудочного сока у детей грудного возраста около 5.0, что способствует перевариваю эмульгированного жира молока желудочной липазой. К тому же, есть основания полагать, что при длительном употреблении молока в качестве основного продукта питания у детей грудного возраста имеет место адаптивное усиление синтеза желудочной липазы.
Хотя в желудке взрослого человека не происходит заметного переваривания жиров пище, все же в желудке имеет место частичное разрушение липопротеидных комплексов мембран клеток пищи, что делает жиры более доступными для последующего воздействия на них липазы панкреатического сока. Кроме того, незначительное расщепление диров в желудке приводит к появлению свободных жирных кислот. Последний, поступаю в кишечник, способствуют там эмульгированию жиров (А.Н. Климов и др., 1978).
Расщепление жиров, входящих в состав пищи, происходит у человека и млекопитающих преимущественно в верхних отделах тонкого кишечника, где имеются весьма благоприятные условия для эмульгирования жиров.
После того как химус с некоторой силой как бы впрыскивается в двенадцатиперстную кишку, здесь прежде всего происходит нейтрализация соляной кислоты желудочного сока, попавшей в кишечник пищей, бикарбонатами, содержащимися в панкреатическом и кишечном соках. Выделяющиеся при разложении бикарбонатов пузырьки углекислого газа способствуют хорошему перемешиванию пищевой кашицы с пищеварительными соками. Одновременно начинается эмульгирование жиров. Наиболее мощным эмульгирующим действием на жиры, несомненно, обладают соли желчных кислот.
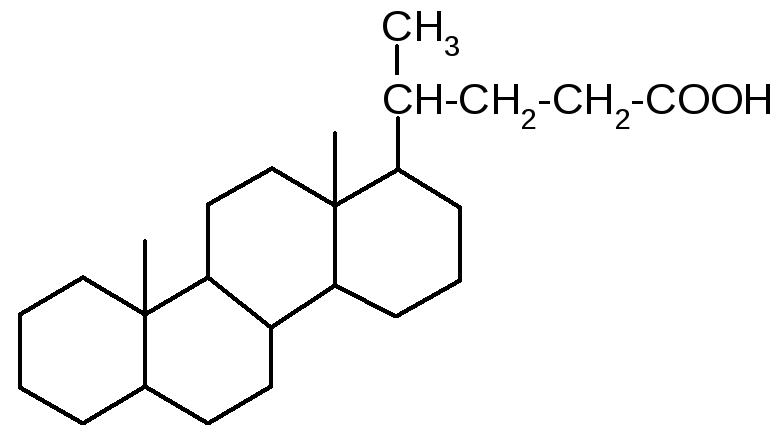 Холановая
кислота
Холановая
кислота
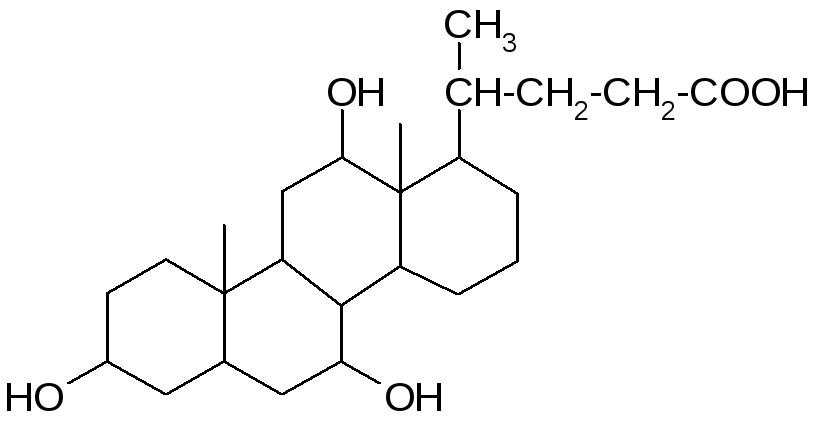
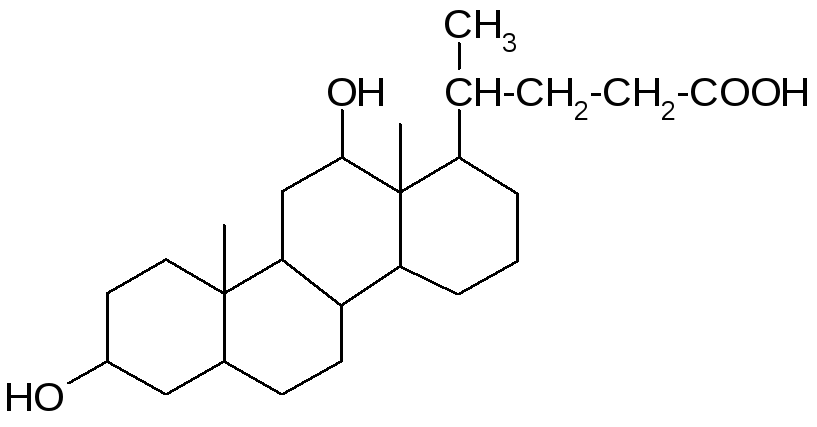
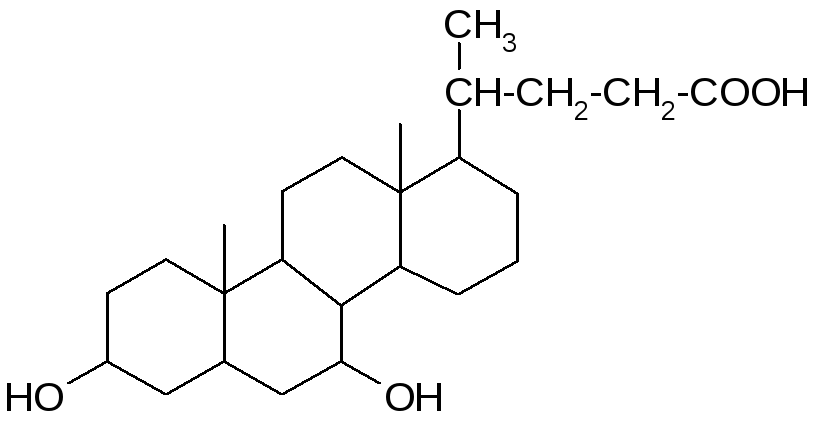
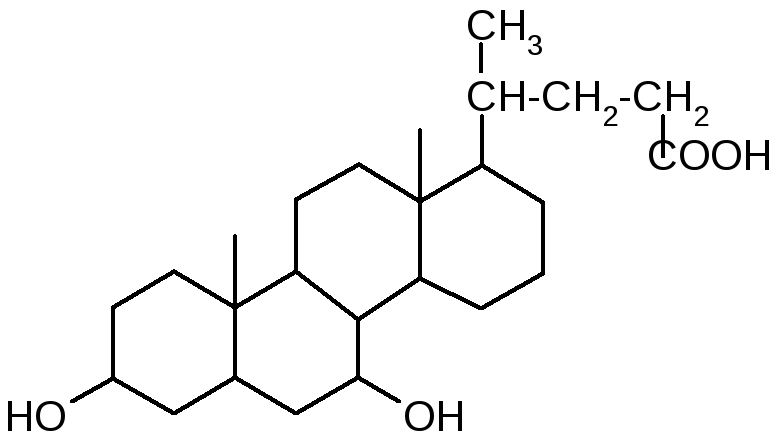
В желчи человека в оновном содержатся: холевая (3, 7, 12 -тригидроксихолановая), дезоксихолевая (3, 12 -дигидроксихолановая) и хенодезоксихолевая (3, 7 -дигидроксихолановая) кислоты.
Холевая кислота Дезоксихолевая кислота
 Хенодоксихолевая
кислота
Хенодоксихолевая
кислота
Кроме того, в желчи человека в малых (следовых) количествах содержится литохолевая (3 - гидроксихолановая) кислота, а также аллохолевая и уреодезоксихолевая кислоты – сереоизомеры холевой и хенодезоксихолевой кислот.
Как уже отмечалось, желчные кислоты присутствуют в желчи в конъюгированной форме, т.е. в виде гликохолевой, гликодезоксихолевой, гликохенодезоксихолевой (около 2/3 – 4/5 всех желчных кислот) или таурохолевой тауродезоксихолевой и таурохенодезоксихолевой (около 1/5 – 1/3 всех желчных кислот). Эти соединения иногда называют парными, так как они состоят из двух компонентов – из желчной кислоты и глицина или из желчной кислоты и таурина.
Заметим, что соотношение между конъюгатами этих двух видов может меняться в зависимости от характера пищи: в случае преобладания в ней углеводов увеличивается относительной содержание глициновых конъюгатов, а при высокобелковой диете – тауриновых конъюгатов (Т.К.Шлыгин, Ф.И.Комаров, 1978). Строение этих конъюгатов может быть представлено в следующем виде.
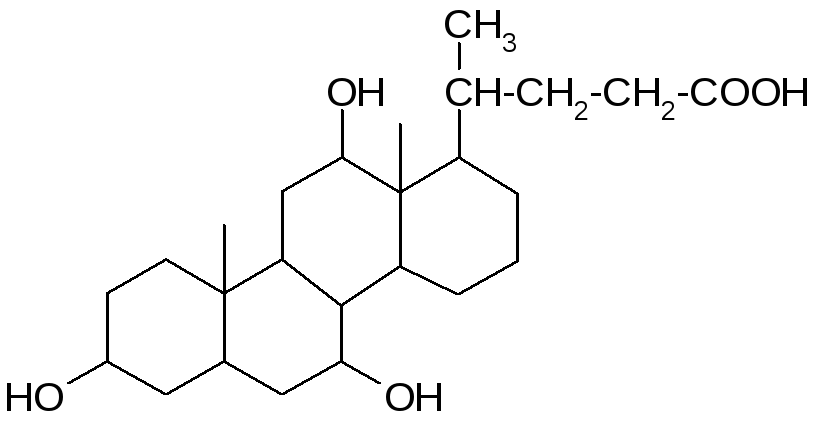 + NH2
–CH2
–COOH
+ NH2
–CH2
–COOH
Холевая кислота глицин (гликокол)
 Гликохолевая
кислота
Гликохолевая
кислота
Хенодезоксихолевая кислота Таурин
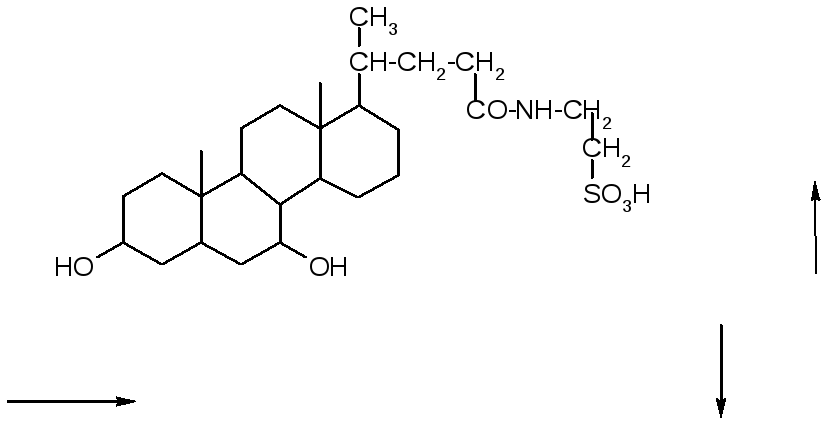
 + NH2
–CH2
–CH2
–SO3H
+ NH2
–CH2
–CH2
–SO3H
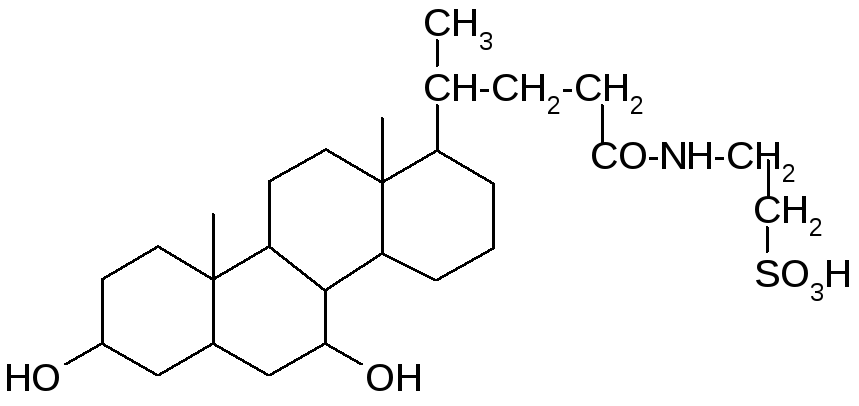 Таурохенодезоксихолевая
кислота
Таурохенодезоксихолевая
кислота
Считается, что только комбинация: соль желчной кислоты + ненасыщенная жирная кислота + моноглицерид способна дать необходимую степень эмульгирования жира. Соли желчных кислот резко уменьшают поверхностное натяжение на поверхности раздела жир/вода, благодаря чему они не только облегчают эмульгирование, но и стабилизируют уже образовавшуюся эмульсию.
Желчные кислоты выполняют также важную роль в качестве своеобразного активатора панкреатической липазы, под влиянием которой происходит расщепление жира в кишечнике. Вырабатываемая в поджелудочной железе липаза расщепляет триглицериды, находящиеся в эмульгированном состоянии. ( действие липазы на триглицериды - слайд №8)Считают, что активирующее влияние желчных кислот на липазу выражается в смещении оптимума действия данного фермента с pH 8.0 до pH 6.0, т.е. до той величины pH, которая более постоянно поддерживается в двенадцатиперстной кишке в ходе переваривания жирной пищи. Конкретный же механизм активации липазы желчными кислотами пока не ясен.
Есть основания считать, что панкреатическая липаза существует двух типов: одна из них специфична в отношении эфирных связей в α–положении триглицерида, а другая – гидролизирует связи в β–положении. Полный гидролиз триглицеридов происходит постадийно: сначало быстро гидролизируются α– и α‘ связи, а потом уже медленно идет гидролиз –моноглицерида (Малер и Кордис, 1970). Однако чаще всего этот гидролиз не происходит и моноацилглицерол непосредственно включается в мицеллу и проникает в энтероцит ворсинки кишечника. В последнее время считают (Ленинджер А., 1985), что поджелудочная железа вырабатывает полипазу, которая в 12ти-перстной кишке совместно с колипазой и солями желчных кислот, образуется активированный комплекс, способствующий гидролизу триацилглицеролов.
Необходимо считать, что в расщеплении жиров участвует также кишечная липаза. Однако, активность ее невысока. К тому же эта липаза катализирует гидролитическое расщепление моноглицеридов и не действует на ди- и триглицериды. Таким образом, практически основными продуктами, образующимися в кишечнике при расщеплении пищевых жиров, являются дирные кислоты, β-моноглицериды и глицерин.
Всасывание жиров в кишечнике. Всасывание происходит в проксимальной части тонкого кишечника. Тонко эмульгированные жиры (величина жировых капелек эмульсии не должна превышать 0.5 мкм) частично могут всасываться через стенку кишечника без предварительного гидролиза. Однако основная часть жира всасывается лишь после расщепления его панкреатической липазой на жирные кислоты, β-моноглицериды и глицерин. Заметим, что в целом в кишечник при распаде жира глицерина образуется немного. Жирные кислоты с короткой углеродной цепью (менее 10 С-атомов) и глицерин, будучи хорошо растворимы в воде, свободно всасываются в кишечнике и поступают в кровь воротной вены, а оттуда в печень, минуя какие-либо превращения в кишечной стенке. Сложнее дело обстоит с жирными кислотами с длинной углеродной цепью и моноглицеридами. Вссывание этих соединений происходит при участии желчи и главным образом желчных кислот (желчные кислоты - слайд №9) , входящих в ее состав (состав желчи – слайд №10). В желчи соли желчных кислот, фосфатиды и холестерин содержатся в соотношении 12.5 : 2.5 : 1.0 (А.Н.Климов и др., 1978). Жирные кислоты с длинной цепью и моноглицериды в просвете кишечника образуют с этими соединениями устойчивые в водной среде мицеллы (мицеллярный раствор) (мицелла – слайд №11) . Структура этих мицелл такова что их гидрофобное ядро (жирные кислоты, глицериды и др.) оказывается окруженным снаружи гидрофильной оболочкой из желчных кислот и фосфатидов. Размеры мицелл примерно в 100 раз меньше самых мелких эмульгированных жировых капель. В составе мицелл высшие жирные кислоты и моноглицериды переносятся с места гидролиза жиров к всасывающей поверхности кишечного эпителия. Относительно механизма всасывания жировых мицелл единого мнения нет. Одни исследователи считают, что в результате так называемой мицеллярной диффузии, а возможно и пиноцитоза, мицеллы целой частицей проникают в эпителиальные клетки ворсинок. Здесь происходит распад жировых мицелл, при этом желчные кислоты сразу же поступают в ток крови и через систему воротной вены попадают в печень, откуда они вновь секретируются в составе желчи. Другие исследователи допускают возможность перехода в клетки ворсинок только липидного компонента жировых мицелл. А соли желчных кислот, выполнив свою физиологическую роль, остаются в просвете кишечника. И лишь потом, в подавляющем своем большинстве, они всасываются в кровь (в подвздошной кишке), попадают в печень, а затем выделяются с желчью. Таким образом, и те и другие исследователи признают, что происходит постоянная циркуляция желчных кислот между печенью и кишечником. Этот процесс получил названия печеночно-кишечной (энтерогепатической) циркуляции.
С помощью метода меченых атомов было показано, что в желчи содержится лишь небольшая часть желчных кислот (10-15 процентов от общего количества), вновь синтезированных печенью. Т.е. основная масса желчных кислот желчи (85-90 процентов) – это желчные кислоты, реабсорбированные в кишечнике и повторно секретируемые в составе желчи. Установлено, что у человека общий пул желчных кислот – примерно 2.8 – 3.5г., при этом совершают они 5-6 оборотов за сутки (Г.К.Шлыгин и Ф.Ф.Комаров, 1978).
IV.(ресинтез триглециридов – слайд №13) Ресинтез жиров в стенке кишечника. В стенке кишечника синтезируются жиры, в значительной степени специфичные для данного вида животного и отличающиеся по своей природе от пищевого жира. В известной мере это обеспечивается тем, что в синтезе триглицеридов (а также фосфатидов) в кишечной стенке принимают участие наряду с экзогенными и эндогенные жирные кислоты. Однако, способность организма к осуществлению в стенке кишечника синтеза жира, специфичного для данного вида животного, все же ограничена. А.Н.Лебедевым показано, что при скармливании животному, особенно предварительно голодавшему, больших количеств чужеродного жира (например, льняного масла или верблюжьего сала) часть его обнаруживается в жировых тканях животного в неизменном виде. Жировые депо, скорее всего, являются единственной тканью, где могут складываться чужеродные жиры. Липиды, входящие в состав протоплазмы клеток других органов и тканей, отличаются высокой специфичностью, их состав и свойства мало зависят от пищевых жиров. Механизм ресинтеза триглицеридов в слетках стенки кишечника в общих чертах сводится к следубщему: первоначально из жирных кислот образуется их активная форма – ацил – Ko, после чего происходит ацилирование моноглицеридов с образованием сначала диглицеридов, а затем триглицеридов.
Моноглицерид + R –CO –S –KoA Диглицерид + HS –KoA
Диглицерид + R1 –CO –S-KoA Триглицерид + HS-KoA
Таким образом, в клетках кишечного эпителия высших животных моноглицериды, образующиеся в кишечнике при переваривании пищи, могут ацилироваться непосредственно без промежуточных стадий.
Однако, в эпителиальных клетках тонкого кишечника содержатся ферменты – моноглицеридлипаза, расщепляющая моноглицерид на глицерин и жирную кислоту, и глицеролкиназа, способная превращать глицерин (образовавшийся из моноглицерида или всосавшийся из кишечника) в альфа-глицерофосфат. Последний, взаимодействуя с активной формой жирной кислоты – ацил-KoA, дает фосфатную кислоту, которая затем используется для ресинтеза триглицеридов и особенно фосфатидов.
V. Переваривание и всасывание фосфатидов и холестерола. Вводимые с пищей фосфатиды подвергаются в кишечнике воздействию специфических гидролитических ферментов, разрывающих эфирные связи между компонентами, входящими в состав фосфолипидов. Принято считать, что в пищеварительном тракте распад фосфатидов происходит при участии фосфолипаз А1, А2, С и Д (или фосфатидаз А, В, С и Д), выделяемых с панкреатическим соком. Ниже приведена схема гидролитического расщепления фосфатидов.
Фосфолипаза А1
Фосфолипаза
А2
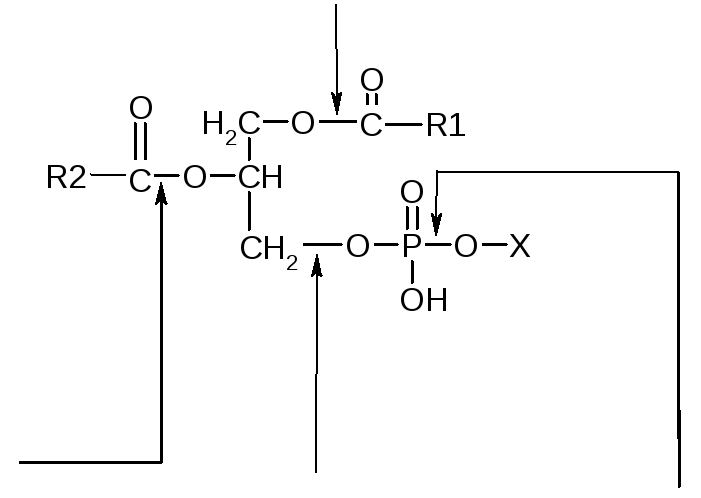
Фосфолипаза C Фосфолипаза D
Лецитин
Схема 23. Точки действия фосфолипаз А, В, С и L при гидролитическом расщеплении лецитина.
В скобках указан шифр ферментов согласно классификации и номенклатуре ферментов.
Схема . Точки действия фосфолипаз А, В, С и D при гидролитическом расщеплении фосфатидов.
R1 – радикал насыщенной жирной кислоты; R2 – радикал ненасыщенной жирной кислоты, Х – остаток азотистого основания (холина и этаноламина). В скобках указан шифр ферментов согласно классификации и номенклатуре ферментов.
Фосфолипаза А2 катализирует гидролитической отщепление жирной кислоты в положении β. Образующийся при этом продукт носит название лизофосфатида. Лизофосфатиды (лизолецитины и лизокефалины) токсичны и вызывают разрушение мембран клеток. В кишечнике, однако, эта опасность устраняется действием на лизофосфатиды фосфолипазы А1, сразу же инактивирующей лизофосфатид путем отщепления жирной кислоты в положении α‘. В результате образуется глицерол -3 –фосфорилхолин. Есть основания считать, что фосфолипаза А1 способна также катализировать отщепление сразу обеих жирных кислот (в положении α‘ и β). Фосфолипаза С вызывает гидролиз связи между фосфорной кислотой и глицерином, а фосфолипаза Д отщепляет Х – группу (продуктом гидролиза при этом оказывается фосфатидная кислота).
Под воздействием всего этого комплекса ферментов фосфатиды расщепляются с образованием глицерина, высших жирных кислот, азотистого основания и фосфорной кислоты.
Необходимо отметить, что подобный механизм расщепления фосфолипидов существует и в тканях организма; катализируется этот процесс тканевыми фосфолипазами. Заметим, что последовательность реакций расщепления фосфатидов на отдельные компоненты еще неизвестна.
Механизм всасывания высших жирных кислот и глицерина был уже рассмотрен, фосфорная кислота всасывается кишечной стенкой главным образом в виде натриевых или калиевых солей; азотистые основании (холин и этаноламин) всасываются в виде своих активных форм.
Как уже отмечалось, в энтероцитах кишечной стенки происходит ресинтез фосфатидов. Необходимые компоненты для синтеза: высшие жирные кислоты, глицерин, фосфорная кислота, органические азотистые основания (холин или этаноламин) поступают в эпителиальные клетки при всасывании из полости кишечника, поскольку они образуются при гидролизе пищевых жиров и липидов; частично эти компоненты доставляются в эпителиальные клетки кишечника с током крови из других тканей. Ресинтез фосфатидов идет через стадию образования фосфатидной кислоты. Подробно синтез фосфолипидов изложен на стр. 307-308.
Что же качается холестерина, то он попадает в пищеварительные органы человека преимущественно с яичным желтком, мясом, печенью, мозгом. В организм взрослого человека ежедневно вводится 0.1 – 0.3 холестерина, содержащегося в пищевых продуктах либо в виде его эфира (холестеридов). Эфиры холестерина расщепляются на холестерин в жирные кислоты при участии особого фермента панкреатического и кишечного соков – холестеразой. Не растворимый в воде холестерин, подобно живым кислотам, всасывается в кишечнике лишь в присутствии желчных кислот.
VI. ( ХМ- хиломикроны, их состав- слайд №14) Образование хиломикронов и транспорт липидов. Ресинтезированные в эпителиальных клетках кишечника триглицериды и фосфатиды, а также поступивший в эти клетки из полости кишечника холестерин (здесь он может частично этерифицироваться) соединяются с небольшим количеством белка и образуют относительно стабильные комплексные частицы – хиломикроны. Последние содержат около 2% белка, 7% фосфатидов, 8% холестерина и его эфиров и свяше 80% триглицеридов. Диаметр хиломикронов колеблется от 100 до 5000 нм. Благодаря большим размера частиц хиломикроны не способны проникать из эндотелиальных клеток кишечника в кровеносные капилляры и диффундируют в лимфатическую систему кишечника, а из последней – в грудной лимфатический проток. Затем из грудного лимфатического протока хиломикроны попадают в кровяное русло, т.е. с помощью хиломикронов осуществляется транспорт экзогенных триглицеридов, холестерина и, частично, фосфатидов из кишечника через лимфатическую систеу в кровь. Уже через 1-2 час после приема пищи, содержащей липиды, наблюдается алиментарная гиперлипемия. Это – физиологическое явление, характеризующееся в первую очередь повышением концентрации триглицеридов в крови и появлением в ней хиломикронов. Пик алиментарной гиперлипемии приходится на 4-6 часов после приема жирной пищи (А.Н.Климов) (плазма «хилезная»). Повышение содержания липидов в крови после принятия пищи тем выше, чем ниже уровень липидов в крови натощак. Обычно через 10-12 часов после приема пищи содержание триглицеридов возвращается к нормальным величинам, а хиломикроны полностью исчезают из кровяного русла.
Известно, жировая ткань играет наиболее существенную роль в дальнейшей судьбе хиломикронов. В клетки жировой ткани хиломикроны не способны (из-за своих размеров) проникать. В связи с этим триглицериды хиломикронов подвергаются гидролизу на поверхности эндотелия капилляров жировой ткани при участии фермента липопротеидлипазы. В результате образуются жирные кислоты и глицерин. Часть жирных кислот проходит внутрь жировых клеток, а часть связывается с альбуминами сыворотки крови и уносится с ее током. Расщепление триглицеридов хиломикронов в кровеносных капиллярах жировой ткани уменьшению их размеров, а также изменению их физико-химических свойств. Конечным органом, где происходит утилизация хиломикронов в процессе обмена веществ, является печень. (Ленинджер А., 1985).