16. Внутриклеточный липолиз
Адреналин и норадреналин увеличивают скорость липолиза в жировой ткани; в результате усиливается мобилизация жирных кислот из жирового депо и повышается содержание неэстерифицированных жирных кислот в плазме крови. ( внутриклеточный липолиз-слайд №15 ) Адреналин стимулирует через аденилатциклазу синтез цАМФ. В свою очередь цАМФ активирует соответственную протеинкиназу, которая способствует фосфорилированию триглицеридлипазы, т. е. образованию её активной формы. Активная триглицеридлипаза ращепляет триглецирид на диглицерид и жирную кислоту. Затем при действии ди- и моноглицеридлипаз образуются конечные продукты липолиза – глицерин и свободные жирные кислоты, которые поступают в кровяное русло.
17. Этапы β-окисления жирных кислот
Процесс окисления жирных кислот складывается из следующих основных этапов.( этапы β-окисления жирных кислот – слайд №16 )
Активация жирных кислот. Реакция протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима А и ионов магния. Реакция, в результате которой образуется ацил-КоА, катализируется ферментом ацил-КоА-синтетазой.
Транспорт жирных кислот внутрь митохондрий. Переносчиком активированных жирных кислот с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин. Реакция протекает при участии специфического цитоплазматического фермента карнитин-ацилтрансферазы.
18. Β-окисление жирных кислот с чётным числом атомов углерода
(представлено на слайде №17)
Процесс окисления жирной кислоты в митохондриях клетки включает несколько энзиматических реакций.
Первая стадия дегидрирования. Ацетил-КоА в митохондриях подвергается ферментативному дегидрированию, при этом Ацетил-КоА теряет 2 атома водорода в α и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты.
Стадия гидратации. Ненасыщенный ацил-КоА при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА.
Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА дегидрируется. Эту реакцию катализируют НАД –зависимые дегидрогеназы.
Тиолазная реакция. Представляет собой ращепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результе образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой. Образовавшийся ацетил-КоА подвергается окислению в цикле трикарбоновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА, который в свою очередь окисляется до 2 молекул ацетил-КоА.
(этапы β-окисления от бутирил-КоА до ацетил-КоА представлены на слайде №18).
19. Β-окисление жирных кислот с нечётным числом атомов углерода
(представлено на слайде №19)
Жирные кислоты с нечётным числом углеродных атомов окисляются таким же образом, как и жирные кислоты с чётным числом углеродных атомов, с той лишь разницей, что на последнем этапе ращепления образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не две молекулы ацетил-КоА.
20. Обмен глицерина
(общая схема представлена на слайде №20).
21. Метаболизм кетоновых тел.
Кетоновые тела представлены:
- ацетоуксусной кислотой
![]()
- β-оксимасляной кислотой (D-3-гидроксибутират)
![]()
- ацетоном
![]()
Кетоновые тела образуются в печени. В здоровом организме ацетон в крови присутствует в крайне низких концентрациях, образуется в результате спонтанного декарбоксилирования ацетоацетата.
( метаболизм кетоновых тел представлен на слайде №21).
- Первый путь синтеза кетоновых тел. На первом этапе из 2 молекул ацетил-КоА образуется ацетоацетил-КоА. Реакция катализируется ферментом ацетил-КоА-ацетилтрансферазой. Затем ацетоацетил-КоА взаимодействует ещё с одной молекулой ацетил-КоА. Реакция протекает под влиянием фермента гидроксиметилглутарил-КоА-синтетазы. Образовавшийся β-окси-β-метилглутарил-КоА способен под действием гидроксиметилглутарил-КоА- лиазы ращепляться на ацетоацетат и ацетил- КоА.
Ацетоацетат восстанавливается при участии НАД-зависимой D-3-гидроксибутиратдегидрогеназы, при этом образуется D-β-оксимасляная кислота.
-Второй путь синтеза кетоновых тел. Образовавшийся путём конденсации 2 молекул ацетил-КоА ацетоацетил-КоА способен отщеплять коэнзим А и превращаться в ацетоацетат. Этот процесс катализируется ферментом ацетоацетил-КоА-гидролазой.
Известно, что в периферических тканях 3-гидроксибутират способен окисляться до ацетоацетата, а последний активируется с образованием соответствующего КоА-эфира. Ацетоацетат может быть активирован путём переноса КоА с сукцинил-КоА в реакции, катализируемой спецефической КоА-трансферазой. Образовавшийся ацетоацетил-КоА далее расщепляется тиолазой с образованием 2 молекул ацетил-КоА, которые затем включаются в цикл Кребса.
В крови здорового человека кетоновые тела содержаться в очень небольших концентрациях (в сыворотке крови 0,03-0,2 ммоль/л). При патологических состояниях (у лиц с тяжёлой формой сахарного диабета, при голодании, а также у животных с экспериментальным острым стрептозотоциновым или аллоксановым диабетом) концентрация кетоновых тел в сыворотке крови увеличивается и может достигать 16-20 ммоль/л.
Олеат натрия
Na+
Полярная голова

Неполярный хвост
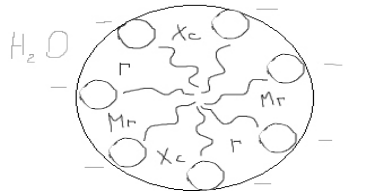
Мицелла олеата натрия
Фиг. II-5. Образование мицеллы мыла в воде. Неполярные хвосты (зигзагообразная линия) молекул олеиновокислого натрия скрыты внутри мицеллы, тогда как отрицательно заряженные карбоксильные группы (кружки) находятся на поверхности мицеллы.
Рис.1.