

СОДЕРЖАНИЕ
ВВЕДЕНИЕ……………………………………………………………….. |
4 |
1. ЭКОЛОГИЯ, ЭВОЛЮЦИЯ И ФЕНЕТИКА НАЗЕМНЫХ БРЮХОНОГИХ МОЛЛЮСКОВ СЕМЕЙСТВ HELICIDAE И HYGROMIIDAE…………………………………………………………................. |
8 |
1.1 Современный ареал надсемейства Helicoidae................................ |
8 |
1.2 Эволюционные связи семейства Helicidae и Hygromiidae……….. 1.3 Полиморфизм наземных брюхоногих моллюсков ……………… |
12 20 |
1.4 Фенетическая изменчивость семейства Helicidae и Hygromiidae.. |
23 |
1.5 Физико-географические и микроклиматические условия района исследования………………………………………………………………. 2. ОБЪЕКТ ИССЛЕДОВАНИЯ……………………………………….... |
30 35 |
3. ПУНКТЫ СБОРА И МАТЕРИАЛ ИССЛЕДОВАНИЯ................. |
41 |
4. МЕТОДИКА ИССЛЕДОВАНИЯ……………………………………. 4.1 Сбор материала и фиксация……..…………………………………… 4.2 Камеральная обработка……………………………………………….. 4.3 Статистические методы………………………………………………. |
45 45 46 48 |
5. ФЕНЕТИЧЕСКАЯ СТРУКТУРА НЕКОТОРЫХ ВИДОВ НАЗЕМНЫХ БРЮХОНОГИХ МОЛЛЮСКОВ СО СТАБИЛЬНЫМ И ФОРМИРУЮЩИМСЯ АРЕАЛОМ В БУГСКО-ПОЛЕССКОМ РЕГИОНЕ……………………………………………………………………… 5.1 Фенетическая структура формирующихся периферийных популяций Cepaea nemoralis L……………………………………………….. 5.1.1 Пространственная изменчивость фенетических параметров в пределах формирующегося ареала Cepaea nemoralis L.……………………… |
51
51
51 |
5.1.2 Хронологическая изменчивость фенетических параметров формирующихся популяций Cepaea nemoralis L………………………….. …… |
57 |
5.1.3 Изменчивость морфометрических параметров в формирующихся периферийных популяциях Cepaea nemoralis L.………………………... |
61 |
5.2 Фенетическая структура формирующихся периферийных популяций Helicella candicans L.Pfr..………………………………………… 5.2.1. Пространственная изменчивость фенетических параметров в пределах формирующегося ареала Helicella candicans L.Pfr……………....... |
66
66 |
5.2.2 Хронологическая изменчивость фенетических параметров формирующихся популяций Helicella candicans L.Pfr.………………………... |
69 |
5.3 Фенетическая структура периферийных популяций Helix pomatia L.в стабильной части ареала………………………………………… 5.3.1 Пространственная изменчивость фенетических параметров периферийных популяций Helix pomatia L. в стабильной части ареала............ |
71
71 |
5.3.2 Хронологическая изменчивость фенетических параметров периферийных популяций Helix pomatia L. в стабильной части ареала……….. 5.3.3 Изменчивость морфометрических параметров периферийных популяций Helix pomatia L.в стабильной части ареала……………………. 6. ФЕНОГЕОГРАФИЯ ПОЛИМОРФИЗМА ПЕРИФЕРИЙНЫХ ПОПУЛЯЦИЙ CEPAEA NEMORALIS L., HELICELLA CANDICANS L. PFR. И HELIX POMATIA L. БУГСКО-ПОЛЕССКОГО РЕГИОНА………………………………………………………………….. ЗАКЛЮЧЕНИЕ…………………………………………………………… СПИСОК ЛИТЕРАТУРЫ……………………………………………….. ПРИЛОЖЕНИЕ……………………………………………………………. |
75
78
82 93 95 99
|
ВВЕДЕНИЕ
За последние десятилетия в изучении природных популяций наметились две основные тенденции. С одной стороны, быстро развились такие специальные направления исследований, как популяционная генетика и популяционная экология; также сформировалась популяционная морфология; в этой отрасли проводится множество исследований. С другой стороны, активно формируется новый общий подход, выражающийся в развитии фенетики популяций. Фенетика популяций – междисциплинарное направление в популяционной биологии, сущностью которого является распространение генетических подходов и принципов на формы, собственно генетическое изучение которых затруднено или невозможно. Предмет данной науки – внутривидовая изменчивость, доводимая в конечном итоге до рассмотрения дискретных, нередко альтернативных вариаций признаков – фенов [1].
Но несмотря на огромные успехи в изучении популяций, от современного знания основных эволюционных факторов и процессов, протекающих на микроэволюционном уровне, до перехода к «управляемой эволюции» (как главной задаче, которую ставит развивающееся человеческое общество перед биологией) еще очень далеко. Одна из главных причин такого положения – недостаточное знание особенностей элементарных эволюционных единиц – популяций. Это положение связано с тем, что до сих пор точному микроэволюционному исследованию были доступны лишь виды, в той или иной степени изученные генетически. Однако подавляющее большинство видов в природе ни сейчас, ни в будущем не смогут быть объектами генетических исследований [2].
Поэтому именно фенетический подход позволит вести исследование природных популяций как бы с генетической точки зрения, но опираясь на материал морфо-физиологического, экологического, этологического исследований на неограниченном спектре видов животного мира.
В данной работе проводится обобщение и систематизация тех знаний, которые получены за многие года изучения изменчивости организмов. Объектом для настоящего исследования выбраны наземные моллюски: Cepaea nemoralis, Helicella candicans, Helix pomatia так как это удобный объект, потому что они характеризуются массовостью, у них слабая подвижность, популяции располагается на незначительных по размеру территориях и на небольшом по площади пространстве. Есть вероятность сделать выборки различных популяций, а также они приурочены к определенным микроклиматическим условиям. Ряд видов моллюсков являются полиморфными. В последнее время привлекают внимание множество существующих споров о причинах и факторах возникновения данного процесса, и о том какую роль в эволюции и в приспособлении организмов к условиям окружающей среды играет внутривидовая изменчивость.
Таким образом, изучение морфологической изменчивости, окрасочного полиморфизма и выявление факторов отбора, формирующих фенотипическую структуру популяций наземных моллюсков представляет большой интерес.
Актуальность темы. В течение последних десятилетий среди зоологических исследований в отечественной науке статус приоритетных вновь обрели фаунистические работы. Это связано с проблемами сохранения биологического разнообразия на фоне прогрессирующего разрушения естественных экосистем. Кроме того, степень изученности наземных моллюсков на территории Беларуси остается крайне недостаточным. Это в полной мере касается и Бугско-Полесского региона. В этой связи изучение фауны наземных гастропод и рассмотрение их эколого-биологических особенностей, конхиологической изменчивости и фенетической структуры следует признать весьма актуальными. Важным представляется исследование особенностей функционирования эколого-фаунистических комплексов в современных условиях, как естественных ландшафтов, так и антропогенно-преобразованных [5, 6].
Целью нашего исследования является выявление особенностей фенетической структуры в формирующихся популяциях Cepaea nemoralis L. и Helicella candicans L. и определить направление микроэволюционных процессов.
Для достижения обозначенной цели были поставлены следующие основные задачи:
- изучить фенетическую изменчивость периферийных популяций в формирующихся ареалах Cepaea nemoralis L. и Helicella candicans L;.
- изучить фенетическую изменчивость в популяциях Helix pomatia в стабильной части ареала;
- сравнить фенетическую структуру некоторых наземных брюхоногих моллюсков в формирующихся и стабильных популяциях;
- у видов интенсивно расширяющих ареал определить направление изменчивости и направление расширения ареала;
Научная новизна. Приведены оригинальные данные по фауне наземных моллюсков Бугско-Полесского региона, экологических комплексов наземных гастропод в антропогенно-преобразованных ландшафтах городов. Для исследуемой территории была проведена попытка осуществить феногеографический анализ наземной малакофауны. Для одного вида были уточнены границы ареалов в пределах обследуемого региона.
Апробация результатов работы. Основные результаты и положения были представлены на научных конференциях:
- на заседаниях научно-исследовательской группы «Фен» (2008-2010 гг.);
- на студенческих научных конференциях биологического факультета «НИРС» (2008, 2010 гг.);
- на региональной научно-практической конференции студентов «Состояние природной среды Полесья и сопредельных территорий» 25 марта 2010 г., Брест;
- на региональной студенческой научно-практической конференции «Устойчивое развитие: экологические проблемы» 25 ноября 2010 г., Брест;
- на региональной научной зоологической конференции «Итоги полевого сезона 2010» 11 декабря 2010 г., Брест;
- на II межуниверситетской студенческой научно-практической конференции «Природа, человек, экология» 28 апреля 2011 г., Брест;
Опубликованность результатов работы – 4 статьи:
1. Молодой, Е.Г. Особенности фенетической структуры наземных моллюсков: Cepaea nemoralis, Helicella candicans, Helix pomatia в пределах Полесского физико-географического региона / Е.Г. Молодой // Состояние природной среды Полесья и сопредельных территорий : материалы регион. научн.-практ. конф. студентов, Брест, 25 марта 2010 / Брест. гос. ун-т имени А.С. Пушкина; под общ. ред. Л.Н. Усачёвой. – Брест : БрГУ, 2010 – 56 с.
2. Молодой, Е.Г. Фенотипическая изменчивость наземного моллюска Cepaea nemoralis в пределах урболандшафтов города Бреста / Е.Г. Молодой // Устойчивое развитие: экологические проблемы: материалы регион. научн.-практ. конф. студентов, Брест, 25 ноября 2010 / Брест. Гос. Ун-т имени А.С. Пушкина. – Брест: БрГУ, 2010 – 34 с.
3. Молодой, Е.Г. Динамика изменчивости окраски и полосатости раковины наземного моллюска Cepaea nemoralis в пределах антропогенных ландшафтов города Бреста / Е.Г. Молодой / Итоги полевого сезона 2010: материалы регион. научн. зоологической конференции, Брест, 11 декабря 2010 / Брест. гос. ун-т имени А.С. Пушкина. – Брест: БрГУ, 2010 – 46 с.
4. Молодой, Е.Г. Популяционная структура наземных брюхоногих моллюсков Брестского Полесья по окраске и полосатости раковины/ Е.Г. Молодой / Человек, природа, экология: материалы II межуниверситетской научн.-практ. конф. студентов, Брест, 28 апреля 2011 / Брест. гос. ун-т имени А.С. Пушкина. – Брест: БрГУ, 2010 – 13 с.
Объем и структура работы. Работа изложена на 144 страницах (в том числе – 94 основного текста), состоит из введения, 6 глав, заключения, списка литературы (34 источника, из которых – 4 на иностранных языках), приложения. Работа иллюстрирована 20 рисунками, содержит 24 таблицы.
1. ЭКОЛОГИЯ, ЭВОЛЮЦИЯ И ФЕНЕТИКА НАЗЕМНЫХ БРЮХОНОГИХ МОЛЛЮСКОВ СЕМЕЙСТВ HELICIDAE И HYGROMIIDAE 1.1 Современный ареал надсемейства Helicoidae
Особи любого вида распределяются в пространстве крайне неравномерно: они встречаются только в тех местообитаниях, где находят подходящие для жизни условия. Часть поверхности суши, в пределах которой распространены и проходят полный цикл своего развития особи данного вида (рода, семейства или определенного типа сообщества), называется ареалом .
В пределах ареала особи того или иного вида обычно распределяются по группам разной величины и занимают те его части, где условия в наибольшей степени отвечают требованиям данного вида. Небольшие по численности группы особей одного вида, как правило, существуют непродолжительное время, после чего они обычно погибают в результате резких изменений почвенно-климатических условий, нападения вредителей, возникновения болезней и ряда других причин. Большие группы особей, которые приспособились к условиям конкретного участка ареала, наоборот, сохраняются длительное время (на протяжении бесчисленно многих поколений). Увеличиваясь в численности при благоприятных условиях и уменьшаясь при неблагоприятных, они имеют перспективу к длительному существованию на данной территории. Такие группировки особей одного вида, в течение длительного времени населяющие определенную часть ареала, свободно скрещивающиеся и относительно обособленные от других совокупностей того же вида, называются популяцией [3].
Широко известные представители Helicoidae – яркая группа, имеющая компактный ареал, четко очерченная анатомически и весьма разнообразна конхиологически. Надсемейство Helicoidea в целом распространено в северном полушарии (рисунок 1). Южнее экватора в Южной Америке по Андам до северной части Чили и северо-запада Аргентины спускаются представители Helminthoglyptidae; на востоке Африки – несколько небольших групп неясного систематического положения; на юго-востоке Азии ниже экватора встречаются многочисленные виды нескольких родов Camaenidae (Papuininae).
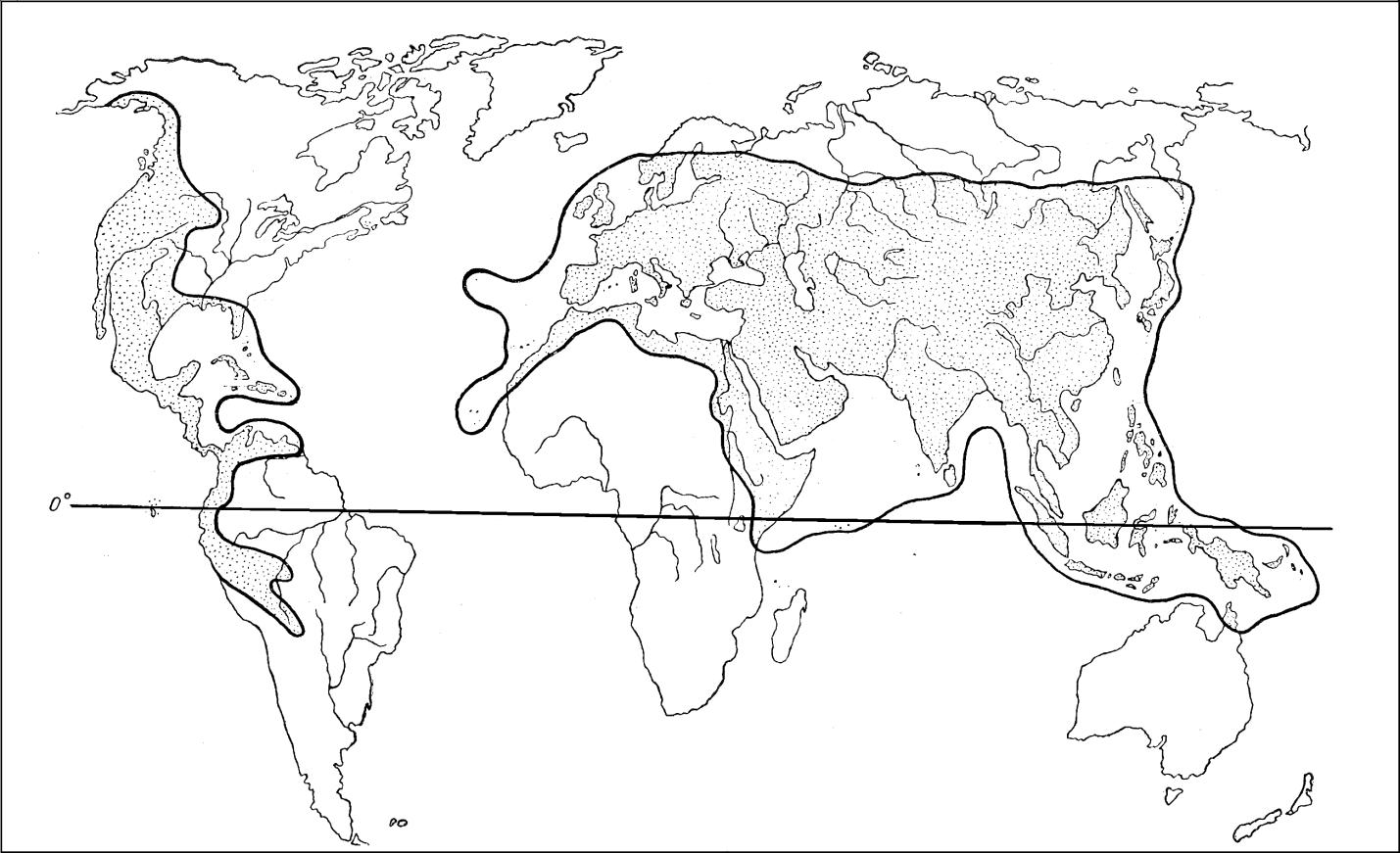
Рисунок 1 – Ареал надсемейства Helicoidea
К этим географическим характеристикам, которые отражают исторически сложившееся распространение, надо добавить, что благодаря вмешательству человека многие виды Helicoidea расселились почти по всему Земному шару.
Такова общая картина распространения основных групп надсемейства.
Исходя из анализа видно, что для Helicoidea фауны нашей страны характерна четкая привязанность подавляющего большинства видов и родов к определенным районам, таксонов с широким распространением очень мало. Это – основная зоогеографическая особенность данной группы: ясное географическое «лицо» большинства таксонов хорошо согласуется с воззрением на филогению группы и историю фауны.
Набор же видов, обитающих на территории нашей страны, сформировался за счет западно-палеарктического зоогеографического центра. За счет ориентального центра сформирована почти вся фауна Helicoidea Дальнего Востока и более половины общего числа видов в фауне Средней Азии. И только два вида восточного происхождения проникли сравнительно недавно на запад, в том числе и на территорию Беларуси (Bradybaenа fruticum и Br. schrencki).
В состав надсемейства Helicoidae входят семь семейств, представители двух из них наиболее распространенны на территории Беларуси: Helicidae и Hygromiidae. В целом на территории Беларуси на сегодняшний момент обитает около 45 видов наземных брюхоногих моллюсков [4].
Широко известные Helicidae – яркая группа, имеющая компактный ареал, четко очерченная анатомически и весьма разнообразна конхиологически. При значительном разнообразии раковин Helicidae отличаются большим постоянством черт внутреннего строения. Даже столь неустойчивый в ряде других семейств признак, как число слизистых желез, здесь жестко закреплен – слизистых желез всегда две, ветвящихся в разной степени – от вообще нерасщепленных до многоветвистых, кустовидных.
Семейство разделяется на два подсемейства – Ariantinae и Helicinae. Распространение семейства в Средиземноморье, причем ареал Ariantinae укладывается целиком внутри ареала Helicinae, занимая преимущественно Южную Европу и не заходя на Пиренейский полуостров, за исключением рода Elona. Но представители последнего встречаются только на склонах Пиренеев и не спускаются ниже предгорий. Наибольшее разнообразие Helicinae наблюдается в двух регионах: во-первых, как и Ariantinae, в Южной и Юго-Восточной Европе и, во-вторых – на территории горной системы Северо-Западной Африки: хребты Большой и Средний Атлас и Антиатлас, причем наиболее своеобразные конхиологические типы свойственны именно этим районам.
Наиболее распространенный на территории Беларуси представитель данного семейства – Cepaea nemoralis L. Основная область его распространения – это преимущественно северные и средние районы Европы. Вид заходит также в Пиренеи и занимает большую часть территории Великобритании. Населяет преимущественно умеренно увлажняемые разреженные леса, опушки, встречается также на влажных лугах, в полосе дюн и в кустарниковых зарослях. В ископаемом состоянии встречаются, начиная с плиоцена [5].
Очень широко распространен в Средней и Юго-Восточной Европе вид Helix pomatia; Также вдоль западных границ на территории Беларуси, Украины, Молдавии и Прибалтики; Завезен в Санкт-Петербург, Москву, Киев. Интродуцирован в Америку. Обитает в лиственных лесах, в зарослях кустарников, часто близ населенных пунктов. Неоднократно найден в плейстоцене и голоцене примерно в пределах современного ареала.
Не менее широко распространенное семейство наземных брюхоногих моллюсков – Hygromiidae. В отличие от Helicidae отличаются большим разнообразием как форм раковин, так и черт внутреннего строения. Число слизистых желез непостоянно – чаще одна, реже две, ветвящиеся в разной степени – от вообще нерасщепленных до многоветвистых, кустовидных.
Семейство разделяется на два подсемейства – Trichiinae и Hygromiinae.
Распространение семейства на большей части Палеарктики, включая Сибирь, Алтай и Дальний восток. Наибольшее число видов – в Южной Европе и на Кавказе. Многие виды завезены в страны Нового Света. В ископаемом состоянии встречаются начиная с верхнего олигоцена (Европа) [5].
Распространение и местообитание Helicella candicans L. – это Средняя и Юго-Восточная Европа с западными районами Украины с Молдавией. Населяет степные участки, иногда встречается по опушкам лесов. Во время летней засухи образует гроздья на стеблях трав. В ископаемом состоянии вид известен начиная с нижнего плиоцена в пределах современного ареала.
Таким образом, современная география надсемейства Helicoidae очень велика. Его представители распространились от таежной зоны умеренного пояса в северном полушарии до влажных экваториальных поясов в южном полушарии.
1.2 Эволюционные связи семейств Helicidae и Hygromiidae
Если не углубляться на анатомических особенностях строения представителей исследуемых семейств, то лучше всего для понимания эволюционных связей проанализировать разработанную А.А. Шилейко филогенетическую схему, основанную на палеонтологическом и морфологическом материале. Рассмотрим эту схему в пространственно-временном аспекте, в частности, насколько она согласуется с данными исторической геологии и палеоклиматологии [5].
Если проанализировать надсемейство в целом, то обратим внимание на такой аспект, что существуют серьезные различия в таксономической структуре трех основных пространственных групп: американской, азиатской и средиземноморской (Рисунок 2). Для первой группы характерны большие роды – до нескольких десятков видов в каждом – при сравнительно малом их числе. В пределах средиземноморской группы картина обратная: здесь имеется множество моно- и олиготипических родов, да и общее их количество намного больше, чем в фауне Нового Света (соответствующие подсчетам: Walden, 1963, а также: Burch, 1969). Исследуемые нами семейства имеют свои корни именно из Средиземноморского участка. Азиатская группа занимает промежуточное положение. Такая разница объясняется неоднозначно (в частности, разной степень расчленения и разъединения горных массивов и совокупностью исторических процессов, приведших к формированию материков в их современном виде), но если принять во внимание ископаемые материалы, то отмеченный факт получает вполне определенное освещение [30].
Судя по палеонтологическим находкам, Helicoidea (не принимая во внимание Sphincterochilidae и Hygromiidae) оформились уже в первой половине мелового периода, поскольку наиболее архаичные группы ныне обитают в северном полушарии на западе Северной Америки, и все ископаемые находки сделаны в том же районе, можно заключить, что местом возникновения надсемейства была юго-западная часть Лавразии, отделенная Тетисом от Гондваны. Между прочим, именно поэтому в южном полушарии Helicoidea первично отсутствуют (Рисунок 3).
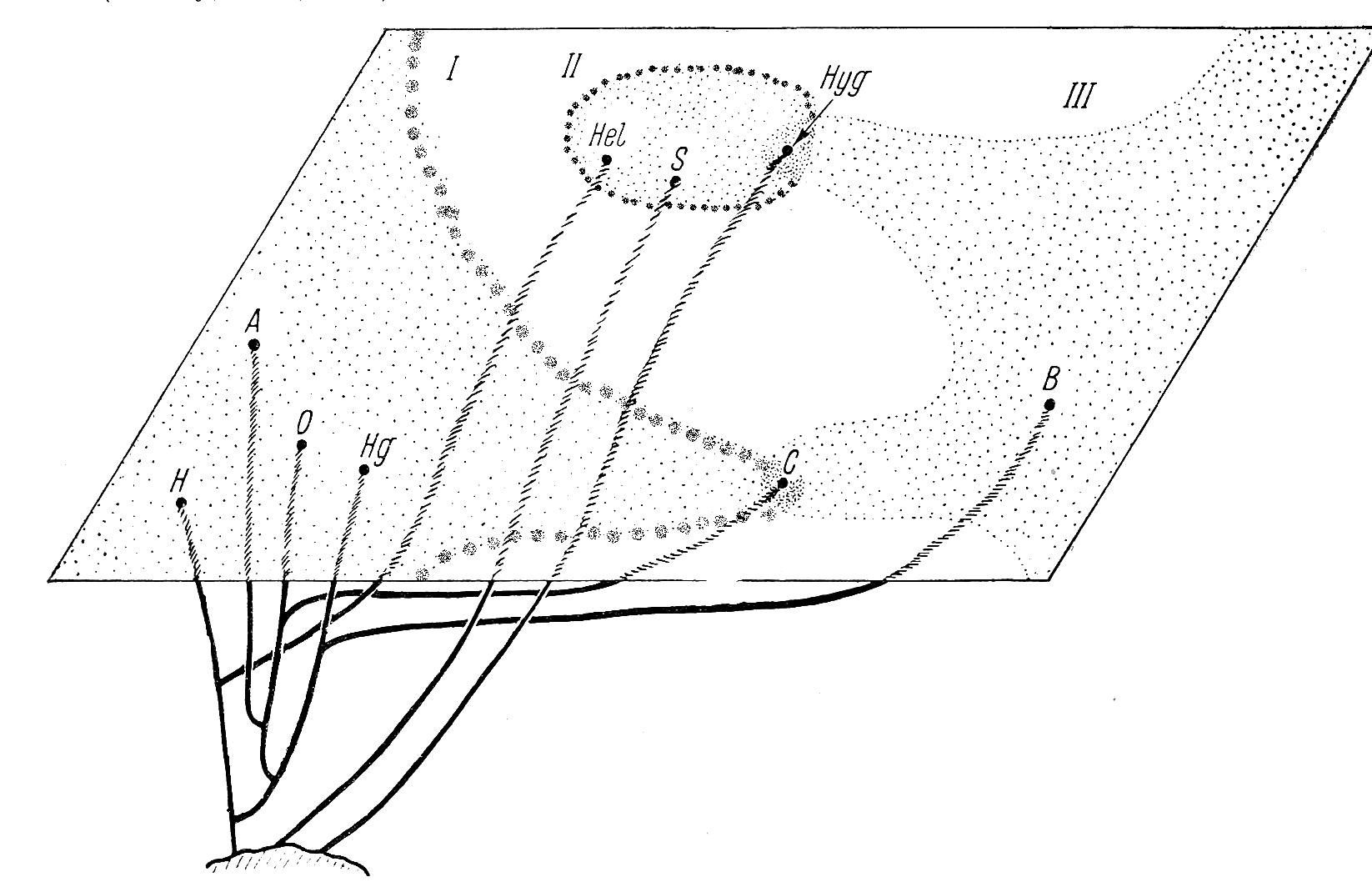
Рисунок 2 – Схема пространственных взаимоотношений между семействами Helicoidea. I – американский участок; II – средиземноморский участок; III – азиатский участок. A – Ammonitellidae; B – Bradibaenidae; C – Camaenidae; H – Humboldtianidae; Hel – Helicidae; Hg – Helminthoglyptidae; Hyg – Hygromiidae; O – Oreohelicidae; S – Sphincterochilidae.
Распространяясь в начале-середине палеогена в восточном направлении, непосредственные предки Humboldtianidae дали начало современным Helicidae, именно Ariantinae, после чего развитие этих групп осуществлялось независимо благодаря возникновению морского барьера между Европой и Северной Америкой.
В то же время, т.е. в эоцене, представители Helminthoglyptidae и Oreohelicidае имели возможность распространяться в западном направлении, на территорию нынешней Юго-Восточной Азии. Азиатскими производными первых стали Bradybaenida, а вторых – Camaenidae. Пилсбри верно наметил пути миграции американской малакофауны в Азию, но считает, что этот процесс происходил намного раньше – еще в мезозое. Возможно, поскольку речь идет о некоторых других группах, Пилсбри прав, но Helicoidea появились в Азии не раньше начала третичного периода.
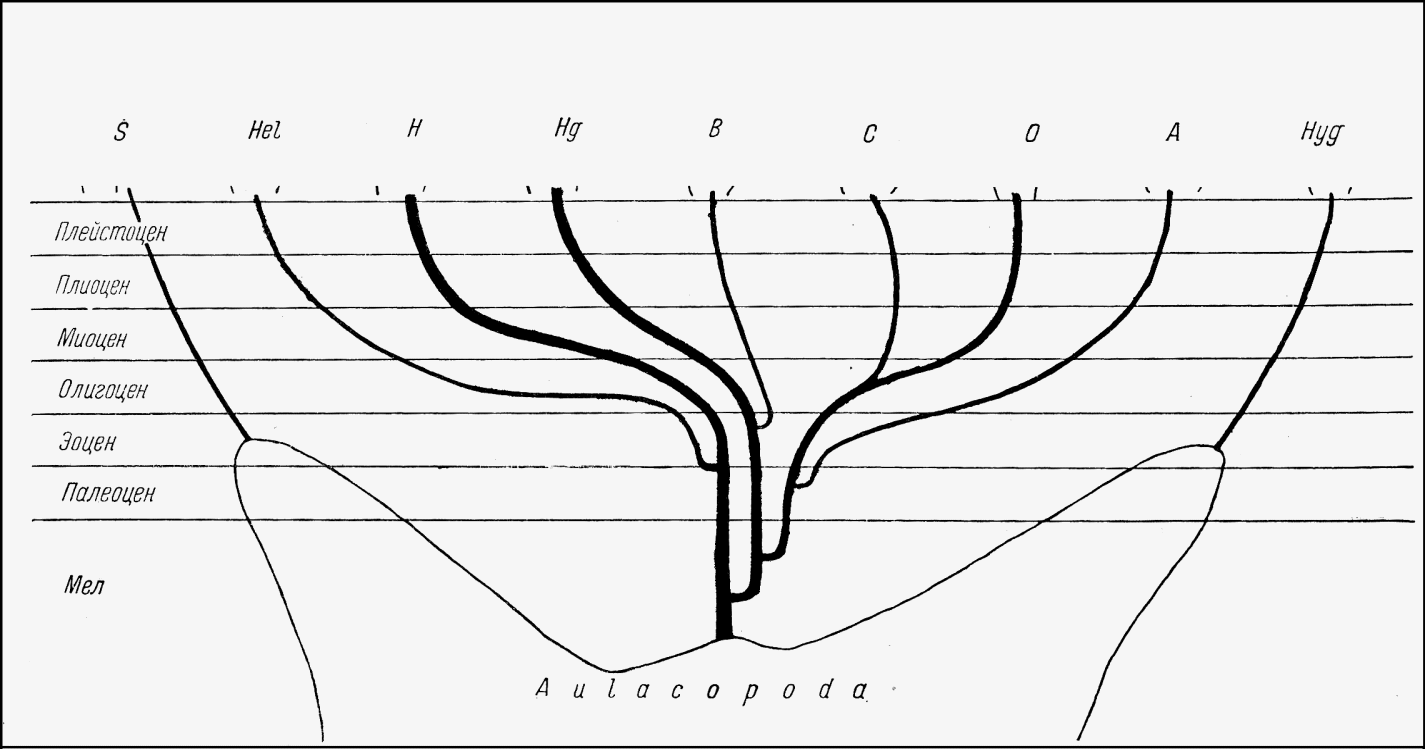
Рисунок 3 – Филогенетическая схема надсемейства Helicoidea. A – Ammonitellidae; B – Bradibaenidae; C – Camaenidae; H – Humboldtianidae; Hel – Helicidae; Hg – Helminthoglyptidae; Hyg – Hygromiidae; O – Oreohelicidae; S – Sphincterochilidae.
Представители семейства Hygromiidae в ископаемом состоянии известны только из Европы, начиная, вероятно, с олигоцена, т.е. с того времени, когда Америка уже окончательно отделилась от Европы; во всяком случае, широкие связи между этими регионами отсутствовал уже с начала палеогена. То же самое относится к Sphincterochilidae и Helicodontidae. Эти факты также свидетельствуют в пользу предположения о независимом возникновении этих групп.
Прямая континентальная связь Европы с Азией открылась лишь в олигоцене-миоцене, и этим объясняется четкое различие в фауне этих районов. Присутствие Hygromiidae в Азии связано с двумя волнами миграций. Первая волна имела мест в течение миоцена – часть Hygromiinae мигрировали из Европы на восток, достигнув берегов Тихого океана (Lindholmomneme), но минуя Среднюю Азию. Позже – в плиоцене-плейстоцене – по линии хребтов Эльбурса и Копетдага в Среднюю Азию проникли ксерофильные Hygromiidae (азиатские Trichiinae проникли сюда тем же путем, но намного раньше – в начале миоцена).
Одновременно со второй волной миграции с запада на восток в обратном направлении продвинулись два вида азиатских Bradybaenidae.
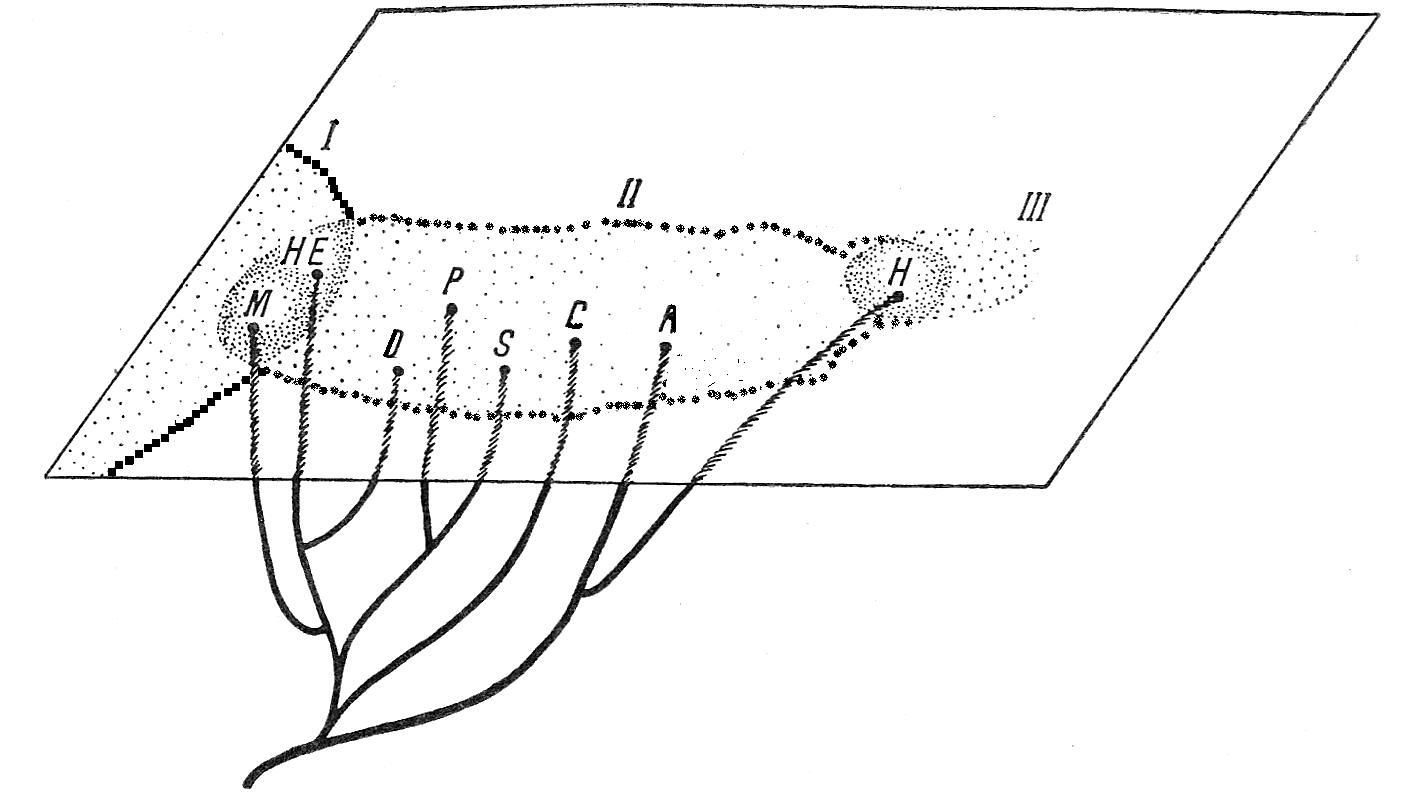
История развития семейства Helicidae вырисовывается, в частности, из сопоставления Ariantinae и Helicinae. Именно представители Ariantinae, известны с эоцена (вымершие роды Galactochilus – верхний эоцен-нижний миоцен, Eurystrophe – верхний эоцен-нижний олигоцен, Klikia – ? средний эоцен-средний плиоцен). Самые ранние находки Helicinae – не раньше среднего миоцена (С. Boettger, Wenz, 1921), причем многие миоценовые группы существуют до сих пор (Theba, Dupotetia, Massylaea, Pseudotachea, Hemicycla, Maurohelix, Caucasotachea, Helix). Показателен следующий подсчет. Если взять все известные таксоны выше вида, имеющие формальную валидность, и, не вдаваясь пока в рассуждения о правомочности их выделения (большое число таксонов свидетельствует как минимум о конхиологическом разнообразии), сравнить число рецентных таксонов с числом вымерших отдельно по подсемействам, получим такие результаты: Ariantinae – рецентных 21, вымерших 12; Helicinae соответственно 49 и 6, т. е. у первых число рецентных таксонов вдвое превышает число вымерших, а у вторых – в 8 раз. Другими словами, можно говорить о расцвете Helicinae сравнительно с Ariantinae в настоящее время (Рисунок 4).
Наибольшее разнообразие Helicinae наблюдается в двух регионах: во-первых, как и Ariantinae, в Южной и Юго-Восточной Европе и, во-вторых – на территории горной системы Северо-Западной Африки: хребты Большой и Средний Атлас и Антиатлас, причем наиболее своеобразные конхиологическпе типы свойственны именно этим районам. Слабая изученность моллюсков Марокко – очень серьезное препятствие для анализа организации высших Helicoidea, поскольку наиболее архаичные формы обитают, по всей вероятности, именно на западе Северной Африки.
Два вымерших рода Ariantinae:
Eurystrophe и Galactochilus

Рисунок 4 – Филогенетические и пространственные взаимоотношения между основными родами Helicidae. I – кавказский участок; II – европейский участок; III – азиатский участок. P – Pseudotachea; НЕ – Hemicycla; Н – Helix; A – Arianta; М – Massylaea; D – Oscarboettgeria; C – Cepaea; S – Succinea.
Заслуживает внимания тот факт, что в Средней и Северной Европе отсутствуют эндемичные Helicinae и вообще это подсемейство здесь представлено тремя родами, имеющими достаточно широкое распространение; в то же время Ariantinae представлены здесь гораздо богаче, причем имеется 12 эндемичных групп, 9 из которых вымерли.
Показателен также удельный вес монотипических групп: у Ariantinae они составляют 35%, у Helicinae 32% всех таксонов, т. е. можно говорить о заметной дискретности и, следовательно, о достаточной древности семейства.
Наконец, необходимо особо подчеркнуть, что решительно все находки ископаемых представителен семейства сделаны в пределах современного ареала, т. е. все частные миграции и перемещения происходили внутри Средиземноморья; есть все основания думать, что заселение Малой Азии и Кавказа произошло не ранее миоцена уже представителями Helicinae; что же касается Ariantinae, то эта группа, вероятно, никогда восточнее Балкан и Карпат не продвигалась.
Признание существования подсемейств Ariantinae и Helicinae диктуется также тем, что в лице этих двух групп мы имеем два типа организации совокупительного аппарата.
Семейство Hygromiidae по сравнению с прочими группами того же ранга отличается наибольшим разнообразием и многоплановостью в строении полового аппарата. В то же время раковина представителей семейства большей частью маловыразительна, причем здесь особенно много случаев конвергентного конхиологического сходства. В пределах семейства намечается несколько достаточно ясных филогенетических линий, реализованных становлением соответствующих подсемейств (рисунок 5).
Здесь хотелось бы остановиться на одном – Trichiinae. Главными в эволюции являются особенности развития женского отдела полового аппарата. Первично заложились 4 равноценные стилофора и дальнейшее развитие сводилось, в частности, к полной или частичной редукции разного их числа либо к видоизменению. Trichiinae и Hygromiinae близкие друг другу, но всегда отличаются одной постоянной особенностью: у первых и у вторых 2 стилофора, расположенных на одной стороне вагины. Необходимо особенно подчеркнуть, что не известно ни одного вида, у которого пара стилофоров на одной стороне вагины была бы развита лучше другой, а такие картины мы неизбежно должны были бы наблюдать, если признать постепенную редукцию стилофоров на одной стороне вагины. Этот факт наталкивает на мысль о возможности мгновенной редукции стилофоров путем мутаций.
Эти рассуждения заставляют принять, что Hygromiinae возникли еще в олигоцене от каких-то древних Trichiinae сначала путем редукции одной пары стилофоров на одной стороне вагины, а затем развивались независимо от Trichiinae.
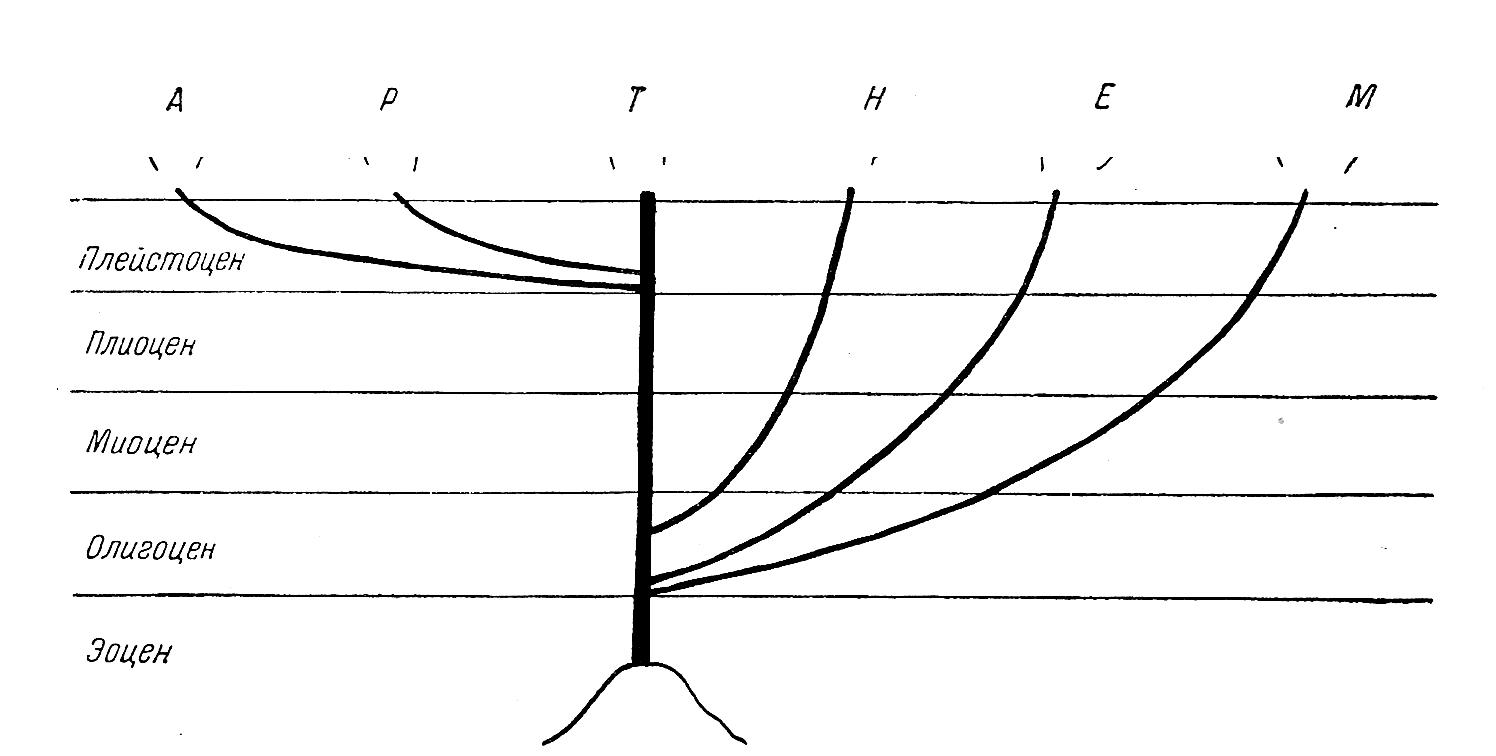
Рисунок 5 – Филогенетическая схема семейства Hygromiidae.
А–Archaicinae; Е–Euomphaliinae; Н–Hygromiinae; М–Metafruticicolinae; Р–Paedhoplitinae; Т–Trichiinae.
Вместе с тем надо обратить внимание, что все азиатские представители семейства – наиболее примитивные представители своих групп.
Анализируя факты, изложенные в систематической части, надо заключить, что развитие анцестральных Trichiinae шло в двух направлениях. Первое направление – восточное: мигрирующие виды продвинулись с территории Эгеиды прямым путем до горных районов Азии, по всей видимости, в начале миоцена. Испытывая воздействие новых условий, эти виды приобрели конхиологический облик, в целом характерный для многих среднеазиатских Helicoidea [5].
Второе направление развития группы нашло выражение в образовании двух группировок – кавказской и европейской, которые и дали начало современным представителям малакофауны данного семейства.
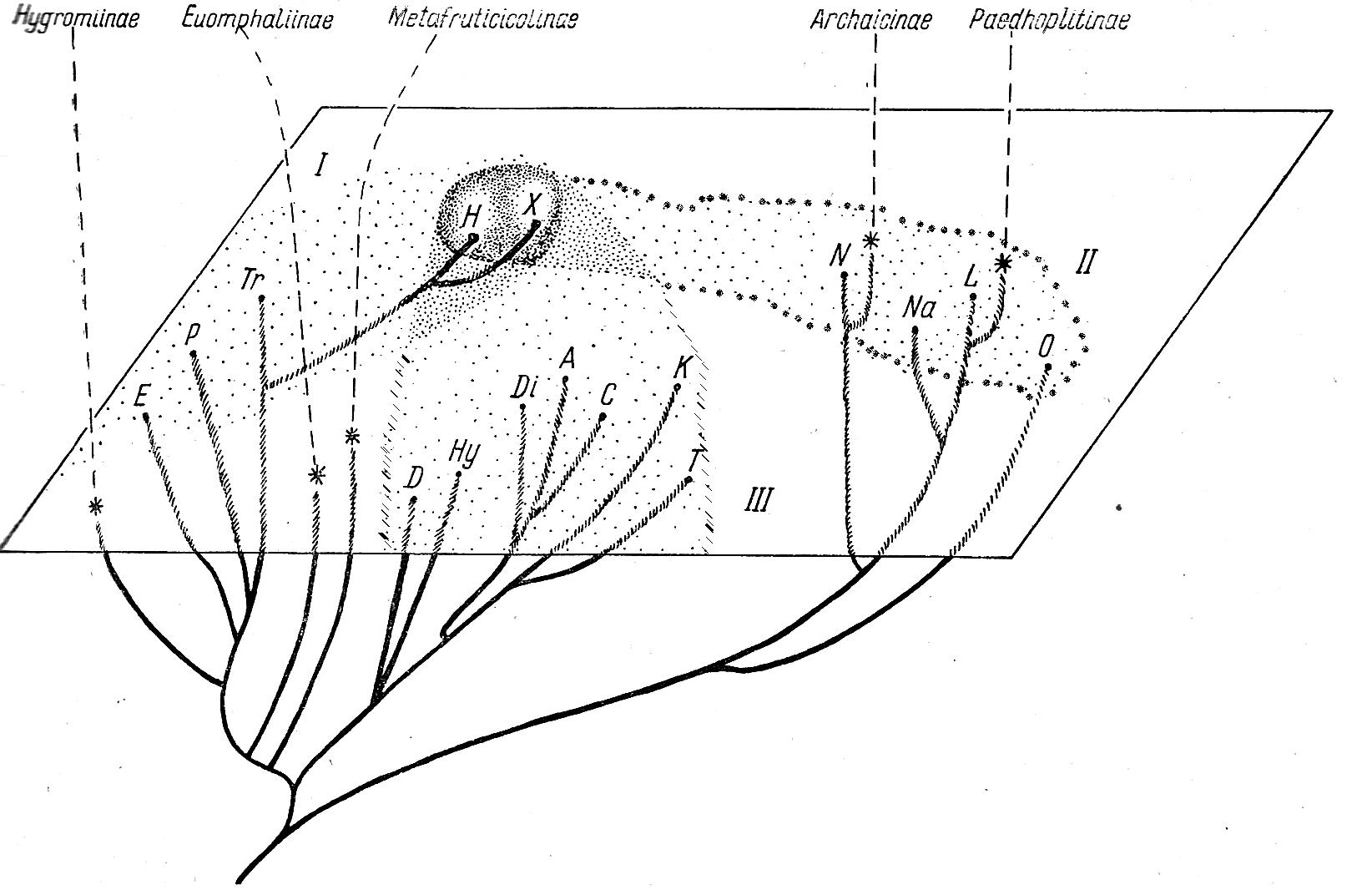
Рисунок 6 – Филогенетические и пространственные взаимоотношения между основными группами Trichiinae и происхождение других подсемейств Hygromiidae. I – европейский участок; II – азиатский участок; III – кавказский участок. А – Anoplitella; С – Caucasigena; D – Diodontella; Di – Dioscuria; E – Edentiella; H – Helicella; Ну – Hygrohelicopsis; К – Kokotschashvilia; L – Leucozonella; N – Nanaja; Na – Narina; О – Odontotrema; P – Plicuteria; T – Teberdinia; Tr – Trichia; X – Xeropicta.
Столь большое место эволюцонисты уделяют деталям строения папиллы пениса в связи с богатством и разнообразием интрапапиллярных структур, эволюция которых есть частное проявление общей эволюции группы. Никакие другие признаки в данном случае не дают столь богатого материала для филогенетических построений.
Очень своеобразная группа Helicella str. Это, по всей видимости, олиготипический род, представители которого обладают парой стилофоров, имеющих в нижней части полости, широко сообщающиеся с полостью вагины, а в верхней части располагается стрела. Напрашивается предположение, что в противоположность остальным Trichiinae здесь прошла редукция не верхней, а нижней (наружной) пары, и отмеченные полости есть остатки нижних стилофоров. В любом случае Helicella достаточно изолированы от прочих родов подсемейства, и в настоящее время нет возможности уверенно говорить об исторических связях [5].
Таким образом, подсемейство Trichiinae, безусловно, монофенетично, но необходимо напомнить, что если европейские и кавказские группы достаточно высоко специализированы, то в Средней Азии до наших дней «законсервировались» примитивные представители, близкие, по всей видимости, к исходным формам, мигрировавшим в Среднюю Азию из Средиземноморья в эоцене-олигоцене; климат Средней Азии не претерпевал резких и внезапных изменений, но наблюдалось очень постепенное и плавное нарастание аридности. Благодаря этому те формы, которые успевали адаптироваться к нарастающей ксерофилизации, оставались в мало измененном состоянии, не имея стимула к бурной эволюционной дивергенции.