

Механизм светового процесса
Свет поглощается хлорофиллом, связанным с белками. Хлорофилл-это производное порфирина, содержащее магний.
Поглощающие свет хлорофилл-белковые комплексы находятся в тилакоидах – мембранных структурах хлоропластов.
При поглощение света хлорофилл переходит в высокоэнергетическое, возбужденное состояние.
В этом состоянии он становится донором высокоэнергетического электрона. Электрон акцептируется электронпереносящей цепью мембраны тилакоидов, а молекула хлорофилла превращается в свободный радикал:

Этот же комплекс хлорофилла с белком осуществляет фоторазложение воды:
![]()
Электроны, образующиеся при восстановлении воды акцептируются радикалами хлорофилла, который возвращается в исходное, невозбужденное состояние.
Электронпереносящая цепь тилакоидных мембран содержит переносчики электронов,которые по механизму действия сходны с переносчиками митохондриальной электронпереносящей цепи. Как и в митохондриях перенос электронов сопровождается синтезом АТФ. Конечным акцептором электронов в тилакоидной мембране служит НАДФ+, который восстанавливается в НАДФН. Акцептором электронов с электропереносящей цепи может быть и радикал хлорофилла. В этом случае АТФ синтезируется, а фотолиз воды и восстановление НАДФ не происходят. С другой стороны возможны фотолиз воды и восстановление НАДФ без синтеза АТФ в электронпереносящей цепи.
Реакции темнового процесса.
Реакция фиксации СО2катализируется рибозо-1,5-бисфосфаткарбоксилазой
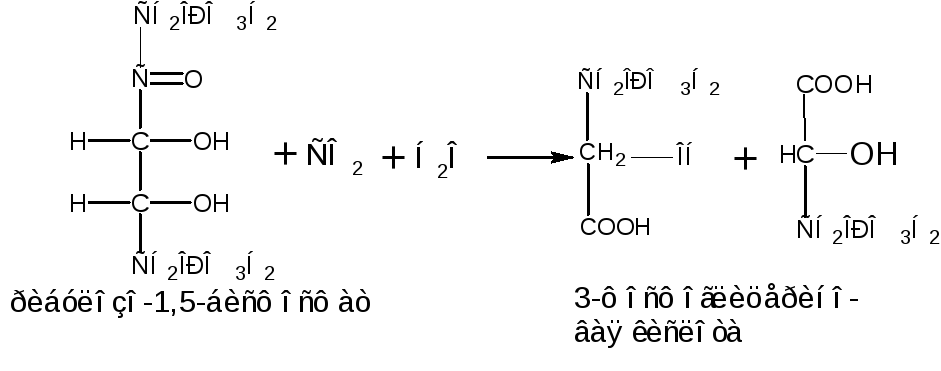
В результате получается две молекулы 3-фосфоглицериновой кислоты. Для синтеза одной молекулы глюкозы необходима фиксация шести молекул СО2.
Далее 3-фосфоглицерат в реакциях, идентичных тем, которые происходят при глюконеогенезе, превращается во фруктозо-6-фосфат:
 Одна
из шести молекул фруктозо-6-фосфата
составляет чистый выход и может
превратиться в глюкозо-6-фосфат. Пять
других молекул фруктозо-6-фосфата
необходимы для возобновления цикла:
они превращаются в шесть молекул
рибулозо-5-фосфата по пути неокислительного
образования пентоз. Наконец,
рибулозо-5-фосфат при участии соответствующей
киназы образует рибулозо-1,5-бисфосфат
и цикл повторяется.
Одна
из шести молекул фруктозо-6-фосфата
составляет чистый выход и может
превратиться в глюкозо-6-фосфат. Пять
других молекул фруктозо-6-фосфата
необходимы для возобновления цикла:
они превращаются в шесть молекул
рибулозо-5-фосфата по пути неокислительного
образования пентоз. Наконец,
рибулозо-5-фосфат при участии соответствующей
киназы образует рибулозо-1,5-бисфосфат
и цикл повторяется.
Следует отметить, что главными нефосфорилированными углеводами, образующимися при фотосинтезе, у большинства растений являются сахароза и крахмал, а не глюкоза.
5.6.3 Биосинтез сахарозы
При синтезе сахарозы донором глюкозы является уридиндифосфатглюкоза (УДФ-глюкоза). Катализирует реакцию сахарозосинтаза:

5.6.4 Биосинтез гликогена и крахмала
Гликоген – это форма запасания углеводов у животных и человека, а крахмал – в клетках растений.
Биосинтез полисахаридов осуществляется путем последовательных реакций трансгликозирования.В реакциях участвует большое число молекул-доноров и молекула акцептора, называемая «затравкой».Донорами биосинтеза служат НДФ-сахара (нуклеозиддифосфатсахара),а затравкой - низкомолекулярный фрагмент синтезируемого полисахарида (п≥4).
Прежде всего глюкоза фосфорилируется при участии фермента гексокиназы:
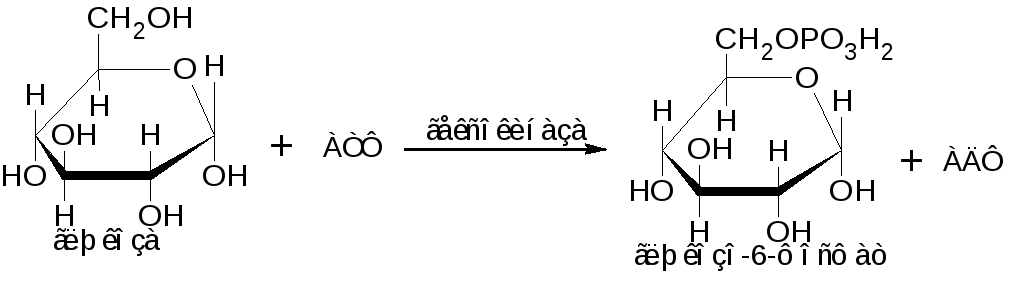
Глюкозо-6-фосфат превращается в глюкозо-1-фосфат:

Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена:

В биосинтезе крахмала может участвовать АДФ-глюкоза. На второй стадии происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы на гликозидную цепь гликогена («затравка»). При этом образуется α-(1→4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозной цепи.
Эта реакция катализируется ферментом гликогенсинтазой (в крахмале-крахмалсинтазой).
 Установлено,
что гликогенсинтаза (крахмалсинтаза)
неспособна катализировать образование
α-(1→6) гликозидной связи, имеющейся в
точках ветвления. Этот процесс катализирует
специальный «ветвящий» фермент-гликозил
(1→6)-трансфераза. Этот фермент катализирует
перенос концевого олигосахаридного
фрагмента, состоящего из 6 или 7 остатков
глюкозы, с нередуцирующего конца одной
из боковых цепей, насчитывающей не менее
11 остатков, на 6-гидроксильную группу
остатка глюкозы той же или другой цепи
гликогена (крахмала).
Установлено,
что гликогенсинтаза (крахмалсинтаза)
неспособна катализировать образование
α-(1→6) гликозидной связи, имеющейся в
точках ветвления. Этот процесс катализирует
специальный «ветвящий» фермент-гликозил
(1→6)-трансфераза. Этот фермент катализирует
перенос концевого олигосахаридного
фрагмента, состоящего из 6 или 7 остатков
глюкозы, с нередуцирующего конца одной
из боковых цепей, насчитывающей не менее
11 остатков, на 6-гидроксильную группу
остатка глюкозы той же или другой цепи
гликогена (крахмала).
Схематически этот процесс можно представить:
