

meditsin1128_1
.pdf2.2. Реологические свойства крови и их влияние на механизм агрегации эритроцитов
Реология — это область механики, которая изучает особенности течения и деформации реальных сплошных сред, одними из представителей которых являются неньютоновские жидкости со структурной вязкостью.
Типичной неньютоновской жидкостью является кровь. Реология крови, или гемореология изучает механические закономерности и особенно изменения физколлоидных свойств крови в процессе циркуляции с различной скоростью и на различных участках сосудистого русла.
Движение крови в организме определяется сократительной способностью сердца, функциональным состоянием кровеносного русла, свойствами самой крови.
При сравнительно малых линейных скоростях течения частицы крови смещаются параллельно друг к другу и оси сосуда. В этом случае поток крови имеет слоистый характер, и такое течение называют ламинарным. Если линейная скорость увеличивается и превышает определенную величину, различную для каждого сосуда, то ламинарное течение превращается в беспорядочное, вихревое, которое называется «турбулентным». Скорость движения крови, при котором ламинарное течение переходит в турбулентное, определяется с помощью числа Рейнольдса, которое для кровеносных сосудов составляет приближенно 1160. Данные о числах Рейнольдса свидетельствуют, что турбулентность возможна лишь в начале аорты и в местах ветвления крупных сосудов. Движение крови по большинству сосудов ламинарно. Кроме линейной и объемной скорости кровотока движение крови по сосуду характеризуется еще двумя важными параметрами, так называемым «напряжением сдвига» и «скоростью сдвига». Напряжение сдвига означает силу, действующую на единицу поверхности сосуда в направлении, тангенциальном к поверхности и измеряется в дин/см2, или в Паскалях. Скорость сдвига измеряется в обратных секундах (с-1) и означает величину градиента скорости движения между параллельно движущимися слоями жидкости на единицу расстояния между ними. Вязкость крови определяется как отношение напряжения сдвига к скорости сдвига, и измеряется в мПа с.
Вязкость цельной крови зависит от скорости сдвига в диапазоне 0,1 — 120 с-1. При скорости сдвига >100 с-1 изменения вязкости не столь выражены, а после достижения скорости сдвига 200 c-1 вязкость крови практически не изменяется. Величину вязкости, измеренную при высокой скорости сдвига (более 120 — 200 с-1), называют асимптотической вязкостью.
Принципиальными факторами, влияющими на вязкость крови, являются гематокрит, свойства плазмы, агрегация и деформируемость клеточных элементов. Учитывая подавляющее большинство эритроцитов по сравнению с лейкоцитами и тромбоцитами, вязкостные свойства крови определяются в основном красными клетками.
Главнейшим фактором, определяющим вязкость крови, является объ-
41
емная концентрация эритроцитов (их содержание и средний объем), называемая гематокритом. Гематокрит, определяемый из пробы крови путем центрифугирования, составляет примерно 0,4 — 0,5 л/л.
Плазма является ньютоновской жидкостью, ее вязкость зависит от температуры и определяется составом белков крови. Более всего на вязкость плазмы влияет фибриноген (вязкость плазмы на 20% выше вязкости сыворотки) и глобулины (особенно Y-глобулины). По мнению некоторых исследователей более важным фактором, ведущим к изменению вязкости плазмы, является не абсолютное количество белков, а их соотношения: альбумин/глобулины, альбумин/фибриноген.
Вязкость крови увеличивается при ее агрегации, что определяет неньютоновское поведение цельной крови, это свойство обусловлено агрегационной способностью эритроцитов. Физиологическая агрегация эритроцитов — процесс обратимый. В здоровом организме непрерывно происходит динамический процесс «агрегация – дезагрегация», и дезагрегация доминирует над агрегацией.
Свойство эритроцитов образовывать агрегаты зависит от гемодинамических, плазменных, электростатических, механических и др. факторов. В настоящее время имеется несколько теорий, объясняющих механизм агрегации эритроцитов. Наиболее известной на сегодняшний день является теория мостикового механизма, согласно которой на поверхности эритроцита адсорбируются мостики из фибриногена или других крупномолекулярных белков, в частности Y-глобулинов, которые при уменьшении сдвиговых сил способствуют агрегации эритроцитов. Чистая сила агрегации является разностью между силой в мостиках, силой электростатического отталкивания отрицательно заряженных эритроцитов и сдвиговой силой, вызывающей дезагрегацию. Механизм фиксации на эритроцитах отрицательно заряженных макромолекул: фибриногена, Y-глобулинов — пока не вполне понятен. Имеется точка зрения, что сцепление молекул происходит за счет слабых водородных связей и дисперсных сил Ван-дер-Ваальса. Существует объяснение агрегации эритроцитов посредством истощения - отсутствия высокомолекулярных белков вблизи эритроцитов, в результате чего появляется «давление взаимодействия», сходное по природе с осмотическим давлением макромолекулярного раствора, что приводит к сближению суспендированных частиц. Кроме этого, существует теория, по которой агрегация эритроцитов вызвана собственно эритроцитарными факторами, которые приводят к уменьшению дзета-потенциала эритроцитов и изменению их формы и метаболизма.
Таким образом, вследствие взаимосвязи между агрегационной способностью эритроцитов и вязкостью крови для оценки реологических свойств крови необходим комплексный анализ этих показателей. Одним из наиболее доступных и широко распространенных методов измерения агрегации эритроцитов является оценка скорости седиментации эритроцитов. Однако в своем традиционном варианте этот тест является малоинформативным, так как не учитывает реологические характеристики крови.
42
2.3.Физическая модель седиментации эритроцитов
Спозиции законов физической химии оседание эритроцитов является своеобразной формой оседания суспензий. Кровь представляет собою с фи- зико-химической точки зрения полидисперсную систему, включающую в себя вещества с различной степенью дисперсности: эритроциты находятся во взвешенном состоянии, белки образуют коллоидный раствор, а некоторые другие органические вещества (мочевина, глюкоза и другие) и соли представляют собою истинный раствор. Оседание эритроцитов, таким образом, происходит в сложной по своему составу дисперсной среде.
Рассмотрение механизма СОЭ может быть понято более детально с точки зрения законов оседания суспензий, причем необходимо учитывать, что оседание суспензий подчиняется законам коагуляции гидрофобных коллоидов.
При этом наиболее существенными моментами являются изменение дисперсности оседающей суспензии и наличие веществ, определяющих нестойкость взвеси. Что касается первого момента, то его выражением при оседании эритроцитов является агломерация последних, которая является, как было изложено выше, наиболее существенным звеном в механизме РОЭ. Что касается второго момента, то из приведенного обзора литературы следует, что наиболее ясную зависимость с оседанием эритроцитов обнаруживают белки крови. Роль белков плазмы вытекает из трех серий наблюдений:
•параллелизм в изменении скорости оседания эритроцитов и содержании отдельных белковых фракций плазмы;
•факт оседания эритроцитов в чистых растворах белков, причем скорость оседания изменяется параллельно изменению концентрации раствора белков;
•клинические наблюдения, свидетельствующие о том, что РОЭ повышена обычно при тех болезненных состояниях, при которых наблюдается накопление в крови грубодисперсных белков.
При этом следует учитывать неодинаковое значение различных белко-
вых фракций плазмы.
Наиболее постоянная зависимость устанавливается между оседанием эритроцитов и фибриногеном плазмы. Известно, что для оседания суспензий (аналогично тому явлению, которое имеет место при коагуляции коллоидов) существенное значение имеют не только свойства вещества, обусловливающего этот процесс (в данном случае фибриногена, являющегося лиофильным коллоидом), но и его количество, что объясняет наблюдающуюся корреляцию между уровнем фибриногена и выраженностью агломерации эритроцитов. Кроме того, известно, что оседание суспензий (также как коагуляция коллоидов) является процессом, протекающим во времени, что объясняет, почему оседание эритроцитов может протекать быстрее и медленнее. Это также связано с уровнем цифр фибриногена в плазме. Известно также, что в дефибринированной крови оседание эритроцитов почти не на-
43
блюдается. Таким образом, очевидно, фибриноген является основным веществом, приводящим взвесь эритроцитов в неустойчивое состояние и обусловливающим ее оседание.
В то же время детальное рассмотрение связи между скоростью оседания эритроцитов и уровнем фибриногена показывает, что не всегда она оказывается прямой и постоянной. Можно наблюдать при этом, что при относительно небольших величинах фибриногена у больного констатируется довольно высокая скорость оседания: оказывается, что в таких случаях повышено количество глобулинов; сочетание, особенно высоких цифр, фибриногена и глобулинов сопровождается очень высокой РОЭ. Важно также подчеркнуть, что оседание эритроцитов может быть более или менее ускорено у больных со значительным уменьшением количества альбуминов, в то же время при его высоких величинах (как это имеет место у здоровых) РОЭ не высока.
Таким образом, глобулиновая и альбуминовая фракции плазмы также принимают участие в течение процесса оседания, причем у альбуминов констатируется тормозящая тенденция, а глобулины проявляют свое действие в сочетании с фибриногеном (в дефибринированной крови лишь очень высокие цифры глобулинов вызывают оседание эритроцитов).
Разнообразная роль различных белковых фракций в процессе оседания также находит свое объяснение при рассмотрении механизма РОЭ с точки зрения законов оседания (коагуляции) и может быть объяснена явлениями коллоидной защиты и сенсибилизации.
Сущность процесса коагуляции состоит в следующем: высокоустойчивые частицы того или иного лиофильного золя адсорбируются на поверхности лиофобных частиц и тем самым предохраняют их от непосредственного соприкосновения одна с другой, а, стало быть, и от агрегации. Коллоидная защита самым тесным образом связана с другим явлением прямо противоположного характера. Оно состоит в том, что прибавление некоторых лиофильных коллоидов к лиофобному не только не увеличивает, но, напротив, понижает порог коагуляции и уменьшает устойчивость этого последнего. Явление это носит название сенсибилизации и заключается в том, что сенсибилизирующий коллоид отнимает от лиофобного адсорбированные на его поверхности стабилизирующие частицы. Поскольку суспензии оседают по законам коагуляции лиофобных коллоидов, все изложенное имеет также прямое отношение к суспензиям.
С этой точки зрения влияние альбуминов на оседание эритроцитов может быть интерпретировано как явление защиты; влияние глобулинов — связано с сенсибилизацией: по-видимому, глобулины сенсибилизируют взвесь эритроцитов к влиянию фибриногена. Можно полагать, что при очень больших количествах глобулины также могут выступать в качестве фактора, приводящего взвесь эритроцитов в неустойчивое состояние, что может объяснить те сравнительно редкие данные эксперимента, когда в дефибринированной крови можно наблюдать оседание эритроцитов.
Как следует из изложенного, в основе реакции оседания эритроцитов
44
лежит явление оседания взвеси эритроцитов в плазме. Оседание суспензий является процессом, происходящим во времени; отмечая величину РОЭ, фактически учитывают степень неустойчивости взвеси эритроцитов, сопровождающуюся более или менее выраженной скоростью ее оседания; это происходит под влиянием лиофильных коллоидов плазмы: фибриногена, глобулинов, альбуминов, которые в естественных условиях в большем или меньшем количестве имеются в плазме. Более выраженная устойчивость взвеси эритроцитов обусловлена постоянным присутствием в плазме альбуминов, в то время как с фибриногеном и глобулинами связана ее неустойчивость. Таким образом, может быть объяснен механизм реакции оседания эритроцитов.
Изложенная физико-химическая теория оседания эритроцитов, не претендуя на исчерпывающую полноту, позволяет, с одной стороны, объяснить некоторые неясные стороны явления оседания, а с другой - синтезировать ряд противоречивых данных, имеющихся в литературе по вопросу о механизме реакции оседания эритроцитов.
Считая фибриноген основным астабилизатором взвеси эритроцитов, нельзя ограничивать объяснение механизма оседания только лишь влиянием белков плазмы, считая их роль в явлениях оседания ведущей, поскольку не исключе возможности участия других ингредиентов плазмы в явлениях защиты и сенсибилизации; в частности, можно рассматривать полипептиды как фактор защиты. С точки зрения явлений защиты и сенсибилизации может рассматриваться также роль различных фракций глобулинов, мукопротеидов, мукополисахаридов и других веществ, описываемых в качестве факторов, играющих роль в механизме оседания эритроцитов.
Кровь является естественной сложной дисперсной системой, включающей в себя различные вещества, находящиеся в сложном взаимодействии как между собой, так и с эритроцитами.
С точки зрения предлагаемого объяснения механизму реакции могут быть объединены различные теории оседания, как, например, электрохимическая и теория лабильности коллоидов плазмы. В самом деле, процесс коагуляции (седиментации) представляет собою сложное явление, для течения которого помимо явлений защиты и сенсибилизации, имеет значение ряд дополнительных условий. Известно, например, что седиментация наступает в изоэлектрическом пункте коллоида (правило Гарди); существенным является также влияние электролитов и среды, в которой находятся взаимодействующие вещества (концентрация Н-ионов); велико значение рН и для явлений защиты.
Таким образом, с точки зрения предлагаемого объяснение оседание эритроцитов не является только показателем изменений соотношения белков плазмы, а течение РОЭ определяется всей совокупностью физикохимических изменений крови.
Процесс оседания эритроцитов немонотонен во времени и можно выделить 3 четко выраженных фазы:
В I фазе под действием земного притяжения эритроциты медленно оседают отдельными клетками. Эта фаза короткая.
45
Во II фазе, более длительной, происходит склеивание эритроцитов. Они образуют кучки различной величины, которые оседают уже более быстро. Агломерация эритроцитов является основным феноменом оседания эритроцитов. Без агломерации эритроциты крови здорового человека осели бы за
1ч на 0,2 мм.
ВIII фазе оседание эритроцитов замедляется за счет того, что агломераты располагаются очень густо, что замедляет их дальнейшее оседание.
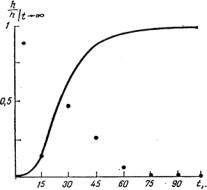
Дробным (через каждые 10 мин) регистрированием опускающегося уровня пограничной зоны показано, что при оседание агрегирующихся эритроцитов человека происходит соответственно кривой, изображенной на рис. 1.5.: вначале, в течение нескольких минут, падение медленное, затем оно ускоряется на 30 - 40 мин, а через 90 - 120 мин граница эритроцитарного столба экспоненциально выходит на практически окончательный уровень.
Иногда обнаруживается S-образность седиментационной кривой h(t): первые 15 — 60 мин опускание границы плазма - эритроциты происходит крайне медленно или не происходит вообще. Вслед за этим наступает весьма быстрая фаза седиментации. «Перекрестное» исследование оседания эритроцитов из такой крови в совместимой плазме (лиц с обычной седиментацией) показало, что фактор, задерживающий начало реакции, содержится в эритроцитах, причем задержка сильно зависит от рН и температуры.
Рис. 1.5. S-образная седиментационная кривая; h—толщина слоя плазмы; кружки — значения hi.
В начале оседания, т. е. за время от одной до нескольких минут, нет четкой границы раздела между чистой плазмой и оседающими эритроцитами. В дальнейшем весь столбик крови в капилляре постепенно разделяется на три зоны (рис. 1.6.):
1. Зона чистой плазмы.
Начинается от верхнего мениска жидкости и доходит внизу до раздела чистой плазмы и оседающих эритроцитов. Над поверхностью раздела имеются отдельные эритроциты (или маленькие агрегаты) в весьма малой концентрации, по-видимому, отмытые от стенок трубки вторично после прохода верхней границы эритроцитарного столба, а также вынесенные из эритроци-
46

тарной зоны восходящими потоками; чем выше, тем меньше число и размер таких «отставших» агрегатов.
Рис. 1.6. Восходящие и нисходящие потоки в прямой (1) и наклонной (2) седиментационных трубах:
а— зона чистой плазмы, б — зона потоков, в — компактная зона.
2.Зона оседающих эритроцитов.
При малых концентрациях на фоне общего движения вниз существуют нисходящие и восходящие потоки эритроцитов. При больших концентрациях касающиеся друг друга агрегаты создают впечатление единого эритроцитарного остова (сети). При этом вытесняемая плазма пробивает в остове извилистые ходы, по которым поднимается с довольно значительной скоростью вверх, увлекая за собой небольшие агрегаты и отдельные эритроциты.
В результате экспериментов было предложено дополнительное деление зоны оседающих эритроцитов 2 еще на три подзоны:
2-1) Верхняя спокойная подзона. Это самая верхняя, высотой 0,5 — 1 мм часть эритроцитарного столба со случайным расположением клеток, которые оседают независимо друг от друга примерно с одинаковой скоростью.
2-2) Подзона потоков. Наиболее значительная часть эритроцитарного столба, которая включает потоки эритроцитов, движущиеся вверх и вниз и возникающие из предыдущей зоны, то ускоряясь, то замедляясь (рис. 3) и разворачиваясь вверх. Обычно бывает не более двух потоков в каждом направлении, а вдоль стенок таких потоков не наблюдается. В этой подзоне, в слое толщиной 50 мкм, эритроциты оседали много медленнее, чем в центре зоны.
2-З) Нижняя спокойная подзона. Расположена тотчас над компактной зоной 3. Размеры - как и в верхней спокойной подзоне.
3.Компактная зона.
Вэтой нижней зоне трубки все эритроциты касаются один другого, движение практически отсутствует, концентрация их максимальна.
Расположение восходящих потоков в вертикальной трубке имеет случайный характер. В наклоненных капиллярах восходящие потоки вполне регулярны (рис. 3) и ускоряют оседание: уже в первые минуты кровь разделяется на слой плазмы и слой эритроцитов по всей длине трубки, и оседание представляет собой сочетание дальнейшего расслоения эритроцитов и плаз-
47
мы, со всплыванием чистой плазмы вверх и опускания столба эритроцитов вниз.
Одиночные эритроциты человека во время оседания вращаются так, что смена способа движения (ребром вниз, наклонно или плоско) происходит приблизительно три раза в минуту. Больше половины времени падения эритроцит движется ребром вниз и скорость такого оседания (0,99 мкм/с) больше, чем скорость при движении наклонно (0,87) или плоской поверхностью вниз (0,76). Средняя скорость оседания одиночного эритроцита в физиологическом растворе (вязкость ≈ 10-3 Па с, плотность ≈ 10-3 кг/м3) равна 0,9, хотя разброс за счет различий в диаметрах эритроцитов, их плотностях и способах падения велик: от 0,59 до 1,20 мкм/с.
Опускание эритроцитов (и агрегатов) сопровождается вытеснением плазмы вверх, т. е. оседание всегда сопровождается возникновением течения. Независимо от того, принимает ли оно форму крупномасштабных восходящих и нисходящих потоков, это течение сопровождается вращением и флуктуациями эритроцитов. Сдвиговый характер течения в крупных потоках и мелкомасштабные движения могут вторично повлиять на оседание, а именно - ускорить его по мере усиления агрегации эритроцитов. Другими словами, агрегация, усиливающая начальное оседание, должна приводить к ускорению возникающих микротечений, а поэтому – к дальнейшему усилению агрегации.
Таким образом, и экспериментальные исследования, и физикохимическая теория седиментации эритроцитов показывают, что центральным движущим звеном процесса оседания эритроцитов является агломерация эритороцитов под влиянием физико-химических факторов изменения крови.
2.4. Математическая модель седиментации эритроцитов в капилляре
Основой теоретического анализа процесса оседания эритроцитов служит обобщенная формула Стокса в виде:
v p − v f = (ρp − ρ f ) g |
kϖ 2 / 3 |
(1.38) |
|
η |
|||
|
|
||
|
f |
|
где vp(x,t) и vf(x,t) - скорости эритроцитов и плазмы в данный момент времени в данном сечении седиментационной трубки;
ρp и ρf - истинные плотности эритроцитов и плазмы; η - вязкость плазмы;
k - коэффициент, зависящий от среднего размера и формы эритроцитарных агрегатов и их объемной концентрации H, характеризующий взвесь в сечении x в момент времени t;
ω - средний объем одного агрегата; g - ускорение силы тяжести. Формула (1.38) представляет собой следствие закона сохранения коли-
чества движения и имеет смысл условие приближенной равномерности (квазистационарности) движения. Формула (1.38) превращается в формулу Стокса для сферической частицы, падающей в безграничной среде, когда vf = 0 и
48
k = (6π)-1 (4π/3)1/3.
Соотношение (1.38) содержит неизвестные функции от x, t: скорости vp(x,t) и vf(x,t), H и N = H/ω, где N – числовая концентрация агрегатов. Связь между vp(x,t) и vf(x,t) дается законом сохранения массы смеси, т.е. в данном случае формулой
v p H + v f (1− H ) = 0 и поэтому: |
|
|
v p = (ρp − ρ f )g kH 2 / 3 (1− H ) |
, |
(1.39) |
ηf N 2 / 3 |
|
|
С позиции общей теории оседание (как относительное движение фаз в смеси) представляет собой диффузию, вызванную несобственным беспорядочным движением частиц, а действующей на них внешней силой.
Формула (1.39) есть определяющее соотношение для двухфазной среды, имеющее смысл закона диффузии взвешенных частиц.
Данная теоретическая модель седиментации эритроцитов достаточно сложна и не имеет практической реализации. Кроме этого, произвольность выбора некоторых коэффициентов при числе оптимизируемых параметров более двух.
Поэтому представляется целесообразным:
-во-первых, максимально упростить формализм математической модели;
-во-вторых, заложить в модель реальные механические свойства эритроци-
та (форму, объем, площадь поверхности, мембраны, деформируемость). Диапазон практической применимости в этом случае существенно рас-
ширяется.
С этой точки зрения интересной является модель, предложенная И.В.Ямайкиной и Э.В.Ивашкевичем.
Математическая модель седиментации эритроцитов построена при исходных допущениях:
1.Оседание происходит «единым фронтом», без деления на зоны;
2.Концентрация эритроцитов в столике постоянна по всей высоте и в каждый данный момент зависит только от длины столбика и исходной концентрации эритроцитов;
3.Оседающие частицы можно считать сферическими;
4.Для каждого отдельного эритроцита или агрегата вся остальная среда представляется сплошной с некоторыми усредненными значениями плот-
ности Δρ(H0) = Δρ(1-H0) и вязкости η(H0) = η(1 + αH02);
5.Эффекты спутанного увлечения плазмы не учитываются.
Рассмотрим случай оседания эритроцитов в гравитационном поле. В ходе
процесса их концентрация в столбике возрастает. В некоторый момент времени t скорость частиц на границе раздела будет выражаться как
|
|
|
H |
0 |
h |
0 |
|
|
|
||
|
|
− |
|
|
|
|
|
||||
|
|
|
|
|
|
|
|||||
|
A 1 |
h(t) |
|
|
|
||||||
v(t) = |
|
|
|
, |
(1.40) |
||||||
1+1− |
αH 0 |
2 h0 |
2 |
||||||||
|
|
|
|||||||||
|
|
h2 (t) |
|
|
|
|
|||||
49
где H0 – объемная концентрация эритроцитов; h(t) – высота столбика оседающих эритроцитов;
H0 – предельная высота столбика осевших эритроцитов; α - реологический параметр;
t – время;
А – скорость падения эритроцита в вязкой жидкости (Re<<1). Коэффициент A рассчитывается по формуле Стокса:
A = |
2gR2 ρ |
, |
(1.41) |
|
9η |
||||
|
|
|
где g = 9,8 м/с – ускорение свободного падения;
R – гидродинамический радиус сопротивления эритроцита или агрегата клетки;
Δρ - разность плотностей оседающей частицы и среды; для эритроцита в плазме Δρ = 50 кг/м3,
для эритроцита в изотоническом буфере (5% раствор цитрата на-
трия pH 7,4)
Δρ = 100 кг/м3
η - вязкость среды.
Длина столбика плазмы при этом равна:
h0 − h(t) = ∫t |
v(τ)dτ. |
(1.42) |
0 |
|
|
Подставим (1.40) в (1.42) и продифференцируем правую и левую части. В результате преобразований получим в левой части функцию высоты столбика эритроцитов, а в правой – линейную функцию времени:
h − h0 + H 0 h0 (1−α) ln |
h − H 0 h0 |
|
− H 0 h0α ln |
h |
= −At |
(1.43) |
||
h |
(1− H |
0 |
) |
h |
||||
0 |
|
0 |
|
|
||||
Рассмотрим предельные случаи.
В начале процесса, при t→0, кинетика оседания выражается линейным уравнением
h = h0 − At |
(1.44) |
При больших временах, зависимость становится экспоненциальной и также хорошо согласуется с опытом:
H = H0 + (1− H0 ) exp(−kt) |
(1.45) |
||
k = |
|
A |
|
H h (1+α) |
|
||
|
0 |
0 |
|
где K – константа скорости оседания.
Из формулы (1.45) видно, что зависимость СОЭ от показателя гематокрита при малых величинах H0 линейна; при увеличении концентрации эритроцитов она становится гиперболической. Это хорошо согласуется с данными опыта.
Как видно из (1.45) начальная СОЭ и константа скорости оседания К пропорциональны квадрату среднего радиуса частицы. При осаждении эрит-
50