

Конспект_лекцій_Заг.вірусологія_0
.pdfчерез плазматичну мембрану. Однак більшість оболонкових і вірусів без оболонки викликають злиття мембран тільки при низькому значенні рН - від 5,0 до 5,75. Якщо до клітин додати слабкі основи (хлорид амонію і ін.), які у ендоцитарних вакуолях підвищують рН до 6,0, злиття мембран не відбувається, вірусні частки залишаються у вакуолях, і інфекційний процес не відбувається. Залежність злиття мембран від значень рН обумовлена конформаційними змінами вірусних білків злиття.
Віруси викликають два типи злиття клітин: 1) злиття зовні і 2) злиття зсередини. Злиття зовні відбувається при високій множинності інфекції і виявляється протягом перших годин після зараження. Навпроти, злиття зсередини відбувається при низькій множинності інфекції, виявляється на порівняно пізніх стадіях інфекційного процесу й обумовлено знову синтезованими вірусними білками. Цей тип злиття викликають ті ж вірусні глікопротеїди, що забезпечують проникнення вірусу в клітину.
Роздягання вірусу в клітині.
Вірусні частки, що проникнули в клітину, повинні роздягнутися для того, щоб викликати інфекційний процес. Суть роздягання полягає у видаленні вірусних захисних оболонок, що перешкоджають експресії вірусного генома. У результаті роздягання звільняється внутрішній компонент вірусу, що здатний викликати інфекційний процес. Роздягання супроводжується рядом характерних рис: у результаті розпаду вірусної частки зникає інфекційна активність, у ряді випадків з'являється чутливість до нуклеаз, виникає стійкість до нейтралізуючого дії антитіл, втрачається фоточутливість при використанні ряду препаратів.
Кінцевими продуктами роздягання є серцевини, нуклеокапсиди чи нуклеїнові кислоти. Для ряду вірусів було доведено, що продуктом роздягання є не голі нуклеїнові кислоти, а нуклеїнові кислоти, зв'язані з внутрішнім вірусним білком. У ряді випадків здатність вірусів викликати інфекційний процес визначається можливістю їх роздягання в клітині даної системи. Тим самим ця стадія є однієї з обмежуючих інфекцію. Роздягання ряду вірусів відбувається в спеціалізованих ділянках усередині клітки (лізосомах, структурах апарата Гольджі, ядерних порах на ядерній мембрані).
Роздягання і внутрішньоклітинний транспорт є взаємозв’язаними процесами: при порушенні правильного транспортування до місць роздягання вірусна частка попадає в лізосому і руйнується лізосомальними ферментами.
Роздягання вірусної частки здійснюється поступово в результаті серії послідовних реакцій. Після руйнування мембран, що оточують піноцитарну вакуоль, нуклеоїди накопичуються в цитоплазмі інфікованих клітин. Через визначений період (приблизно 1 годину) починається наступний етап депротеїнізації, що призводить до повного чи майже повного звільнення нуклеїнової кислоти від білка. У процесі депротеїнізації вірусів можуть брати участь ферменти клітинних лізосом. Початковий період взаємодії вірус —
31
клітина завершується звільненням вірусного геному.
ЛЕКЦІЯ 7. ТРАНСКРИПЦІЯ ВІРУСНИХ ГЕНОМІВ.
Поняття транскрипції Реалізація генетичної інформації у вірусів. Регуляція транскрипції.
Поняття транскрипції
Транскрипція - це переписування інформації з ДНК на РНК за законами генетичного коду. Це означає, що РНК складається з нуклеотидних послідовностей, комплементарних ДНК. Нитки ДНК у ділянці транскрипції розділяються і функціонують як матриці, до яких приєднуються комплементарні нуклеотиди завдяки спарюванню комплементарних основ. Транскрипція здійснюється за допомогою спеціального ферменту ДНКзалежної РНК-полімерази, що зв'язує нуклеотиди шляхом утворення З’- 5’- фосфодіефірних містків. Таке зв'язування відбувається лише в присутності ДНК-матриці.
Продуктами транскрипції в клітині є іРНК. Сама клітинна ДНК, що є носієм генетичної інформацiї не може безпосередньо програмувати синтез білка. Передачу генетичної інформації від ДНК до рибосом здійснює РНКпосредник (іРНК). На цьому заснована центральна догма молекулярної біології, що виражається наступною формулою:
ДНК |
транскрипція |
РНК |
трансляція |
білок, |
|
|
де стрілки показують напрямок переносу генетичної інформації.
Реалізація генетичної інформації у вірусів.
Стратегія вірусного генома у відношенні синтезу іРНК у різних вірусів різна. У ДНК-містких вірусів іРНК синтезується на матриці однієї з ниток ДНК. Формула переносу генетичної інформації в них така ж, як і в клітині:
ДНК |
транскрипція |
РНК |
трансляція |
білок. |
|
|
|
|
|||
ДНК-місткі віруси, |
репродукція |
яких відбувається в ядрі, |
|||
використовують для транскрипції клітинну полімеразу. До цих вірусів відносяться папова-, аденовіруси, віруси герпесу. ДНК-місткі віруси, репродукція яких відбувається в цитоплазмі, не можуть використовувати клітинні ферменти, що знаходяться в ядрі. Транскрипція їх генома здійснюється вірусоспецифічним ферментом – ДНК-полімеразою, що проникає в клітину в складі віріону. До цих вірусів відносяться віруси віспи і ірідовіруси.
РНК-віруси, у яких носієм генетичної інформації є не ДНК, а РНК, вирішують цю проблему особливим чином. У РНК-містких плюс-нитчастих вірусів, у яких функції іРНК виконує сам геном, передача генетичної
32
інформації здійснюється по найбільш простій формулі:
РНК → білок.
До цієї групи вірусів відносяться пікорна-, тога-, коронавіруси. У них немає необхідності в акті транскрипції для синтезу вірусоспецифічних білків. Тому транскрипцію як самостійний процес у цих вірусів не виділяють. Інакше обстоїть справа у вірусів, геном яких не може виконувати функцію іРНК. У клітині синтезується комплементарна геному РНК, що і є інформаційною. Передача генетичної інформації в цих вірусів здійснюється по формулі:
РНК → РНК → білок.
Уцих вірусів транскрипція виділена як самостійний процес в інфекційному циклі. До них відносяться дві групи вірусів тварин:
1)віруси, геном яких представлений односпіральною РНК: ортоміксо-, параміксо-, рабдо-, буньявіруси. Оскільки геномна РНК цих вірусів є мінусниткою, зазначену групу вірусів називають мінус-нитчастими вірусами;
2)віруси, геном яких представлений двоспіральною РНК (диплорнавіруси). Серед вірусів тварин до них відносяться реовіруси.
Уклітині немає ферменту, що може полімеризувати нуклеотиди на матриці РНК. Цю функцію виконує вірусоспецифічний фермент - РНКзалежна РНК-полімераза чи транскриптаза, що знаходиться в складі віріону і разом з ними проникає в клітину.
Серед РНК-містких вірусів хребетних є родина ретровірусів, що мають унікальний шлях передачі генетичної інформації. РНК цих вірусів переписується на ДНК, а ДНК інтегрує з клітинним геномом і в його складі переписується на РНК, яка має інформаційні функції. Шлях передачі генетичної інформації в цьому випадку здійснюється по більш складній формулі:
РНК → ДНК → РНК → білок.
У складі цих вірусів є унікальний вірусоспецифічний фермент, який переписує РНК на ДНК. Цей процес називається зворотною транскрипцією, а фермент - зворотна транскриптаза, чи ревертаза. Той же фермент синтезує нитку ДНК на матриці ДНК. Двоспіральна ДНК після замикання в кільце інтегрує з клітинним геномом, і транскрипцію інтегрованої ДНК у складі клітинних геномів здійснює клітинна ДНКзалежна РНК-полімераза. Оскільки іРНК ретровірусів гомологічна геномній РНК (а не комплементарна їй), ретровіруси є плюс-нитчастими вірусами.
Транскрипція ряду ДНК-містких вірусів — папова-, аденовірусів, вірусів герпесу, парво-, гепаднавірусів — здійснюється в ядрі клітини, і в цьому процесі широко використовуються механізми клітинної транскрипціїферменти транскрипції і модифікації транскриптів. Транскрипція цих вірусів здійснюється клітинною РНК-полімеразою II — ферментом, що здійснює транскрипцію клітинного генома. Однак особлива група транскриптів аденовірусу синтезується за допомогою іншого клітинного ферменту —
33
РНК-полімерази III. У двох інших родин ДНК-містких вірусів тварин (віруси віспи і ірідовірусів) транскрипція відбувається в цитоплазмі. Оскільки в цитоплазмі немає клітинних полімераз, транскрипція цих вірусів має потребу в спеціальному вірусному ферменті - вірусній РНК-полімеразі. Цей фермент
єструктурним вірусним білком.
УРНК-вмісних вірусів транскрипція здійснюється вірусоспецифічними транскриптазами, тобто ферментами, закодованими у вірусному геномі. Ці транскриптази можуть бути як структурними білками, що входять до складу віріону (ендогенна транскриптаза), так і неструктурними білками, що синтезуються в зараженій клітині, але не включаються у віріон.
Синтез комплементарних РНК на батьківських матрицях за допомогою батьківської транскриптази зветься первинною транскрипцією на відміну від вторинної транскрипції, що відбуває на більш пізніх стадіях інфекційного циклу на знову синтезованих, дочірніх матрицях за допомогою нової синтезованої транскриптази. Велика частина іРНК у зараженій клітині є продуктом вторинної транскрипції.
Ускладних РНК-містких вірусів тварин транскрипція відбувається не на матриці голої РНК, а в складі вірусних нуклеокапсидів чи серцевин (транскриптивні комплекси). Зв'язані з геномом капсидні білки не тільки не перешкоджають транскрипції, але і необхідні для неї, забезпечуючи правильну конформацію тяжа РНК, захист його від клітинних протеаз, зв'язок окремих фрагментів генома один з одним, а також регуляцію транскрипції.
Знову синтезовані іРНК виходять із транскриптивних комплексів і транспортуються до рибосом.
Регуляція транскрипції.
Транскрипція вірусного генома строго регулюється протягом інфекційного циклу. Регуляція здійснюється як клітинними, так і вірусоспецифічними механізмами. У деяких вірусів, в основному ДНКмісткими, існує три періоди транскрипції: зверхранній, ранній і пізній. До них відносяться віруси віспи, герпесу, папова-, адено- і ірідовіруси. У результаті зверхранньої і ранньої транскрипції вибірково зчитуються ранні гени з утворенням ранніх іРНК. При пізній транскрипції зчитується інша частина вірусного генома — пізні гени, з утворенням пізніх іРНК. Кількість пізніх генів звичайно перевищує кількість ранніх генів. Більшість ранніх генів є генами для неструктурних білків — ферментів і регуляторів транскрипції — і реплікації вірусного генома. Навпроти, пізні гени здебільшого є генами для структурних білків. Звичайно при пізній транскрипції зчитується весь геном, але з перевагою транскрипції пізніх генів. Фактором регуляції транскрипції в ядерних вірусів є транспорт транскриптів з ядра в цитоплазму, до місця функціонування іРНК — полісомам.
34
ЛЕКЦІЯ 8. СИНТЕЗ ВІРУСНИХ БІЛКІВ
Трансляція інформаційних РНК Загальна інформація про синтез білків
Трансляція інформаційних РНК
Синтез білка в клітині відбувається в результаті трансляції іРНК.
Трансляцією називається процес перекладу генетичної інформації, що міститься в іРНК, на специфічну послідовність амінокислот.
Синтез білка в клітині здійснюється на рибосомах. Процес трансляції складається з трьох фаз:
1) ініціації; 2) елонгації і 3) термінації.
Ініціація трансляції. Ініціація - стадія формування комплексу компонентів, необхідного для дізнавання місця і початку процесу трансляції.
Це найбільш відповідальний етап у процесі трансляції, заснований на дізнаванні рибосомою іРНК і зв'язуванні з її особливими ділянками. Рибосома впізнає іРНК завдяки. «шапочці” на 5’-кінці і сковзає до З’-кінця, поки не досягне ініціативного кодона, з якого і починається трансляція. У еукаріотичній клітині ініціативними кодонами є кодони АУГ чи ГУГ, які кодують валін. З метіоніну починається синтез усіх поліпептидних ланцюгів.
Елонгація трансляції. Елонгація — стадія продовження процесу, що складається з повторюваних дій. Це процес подовження, нарощування поліпептидного ланцюга, заснований на приєднанні нових амінокислот за допомогою, пептидного зв'язку. Відбувається постійне протягання нитки іРНК через рибосому і «декодування» закладеної в ній генетичної інформації. Часто іРНК функціонує одночасно на декількох рибосомах, кожна з яких синтезує ту саму поліпептидну нитку, яка кодується даною іРНК. Група рибосом, що працюють на одній молекулі іРНК, називається полірибосомою, чи полісомою. Розмір полісом значно варіює в залежності від довжини молекули іРНК, а також від відстані між рибосомами. Так, полісоми, що синтезують гемоглобін, складаються з 4-6 рибосом, а високомолекулярні білки синтезуються на полірибосомах, що містять 20 і більше рибосом.
Термінація. Термінація — стадія закінчення процесу трансляції під дією специфічних механізмів. Вона відбувається в той момент, коли рибосома доходить до термінуючого кодона в складі іРНК. Трансляція припиняється, і поліпептидний ланцюг звільняється з полірибосоми. Після закінчення трансляції полірибосоми розпадаються на субодиниці, що можуть увійти до складу нових полірибосом.
Стратегія вірусного генома, що використовує клітинний апарат трансляції, повинна бути спрямована на створення механізму для придушення трансляції власних клітинних іРНК і для виборчої трансляції вірусних іРНК, які завжди знаходяться в значно меншій кількості, чим клітинні матриці. Цей механізм реалізується на рівні специфічного впізнавання малою рибосомальною субодиницею вірусних іРНК, тобто на
35
рівні формування ініціюючого комплексу. Оскільки багато вірусів не придушують синтез клітинних іРНК, у заражених клітинах виникає парадоксальна ситуація: припиняється трансляція величезного фонду функціонально активних клітинних іРНК, і на рибосомах, що звільняються, починається трансляція одиночних молекул вірусних іРНК. Специфічне впізнавання рибосомою вірусних іРНК здійснюється за рахунок вірусоспецифічних ініціативних факторів.
Загальна інформація про синтез білків
Воснові цього синтезу лежить той же механізм, що і при синтезі білка
внормальних клітках. Синтез вірусоспецифічного білка залежить від синтезу вірусної іРНК, але і впливає на нього: якщо синтез білка порушений, відбувається перевиробництво утворених іРНК у місцях її синтезу і гальмується подальший її синтез. Вірусні білки в процесі інфекції синтезуються в надлишковій кількості, чим потрібно для утворення інфекційного вірусу. Наприклад, у клітинах, інфікованих вірусами герпесу, у вірусне потомство включається тільки близько 35% від загальної маси вірусоспецифічних білків, синтезованих у клітинах.
У більшості вірусів синтез білків здійснюється в цитоплазмі; щодо ядерної локалізації синтезу білків деяких вірусів існують сумніви. Відомо, що вірусні білки можуть синтезуватися в одних структурах, а накопичуватися — в інші. Механізми, відповідальні за міграцію вірусних
білків у ядро, не з'ясовані. Відомо лише, що відсутність аргініну в середовищі призводить до придушення міграції структурних білків вірусу герпесу від місця їх синтезу (цитоплазми) до місця зборки віріонів (ядра), хоча синтез ДНК і білка вірусу не порушений. На різних стадіях інфекційного циклу можуть переважно утворюватися то одні, то інші групи вірусоспецифічних білків. Швидкість їх регулюється або на рівні транскрипції (з утворенням іРНК), або на рівні трансляції (зчитування іРНК на рибосомах). У зараженій клітині непропорційно накопичуються іРНК із різних вірусних генів. Механізм цієї непропорційності закладений у самій вірусній частці. Цей же механізм визначає різну ефективність утворення різних білків. Стандартна вірусна частка містить одну молекулу РНК і до 10000 молекул білків. Крім структурних білків, у зараженій клітині синтезуватися і неструктурні білки.
Поряд із синтезом білків у клітині при репродукції вірусу грипу відбувається синтез і вуглеводних ланцюгів, що входять до складу глікопротеїдів. Приєднання вуглеводів здійснюється за допомогою трансфераз, які є клітинними ферментами. Синтез ліпідів також здійснюється клітиною. Вірусна оболонка формується при включенні ліпідів із плазматичної мембрани клітини-хазяїна. Синтез вірусних нуклеїнових кислот і вірусоспецифічних білків відбувається майже одночасно і не менш чим на годину випереджає дозрівання вірусних часток.
36
ЛЕКЦІЯ 9. РЕПЛІКАЦІЯ ВІРУСНИХ ГЕНОМІВ
Реплікація вірусних ДНК Реплікація одноланцюгових вірусних ДНК.
Реплікація односпіральних вірусних РНК Реплікативний цикл +РНК-геномних вірусів.
Реплікація односпіральних вірусних РНК із негативним геномом. Реплікація двоспіральних вірусних РНК.
Реплікація ретровірусів
Реплікація вірусних ДНК.
Механізм реплікації геному ДНК-містких вірусів в основному подібний з механізмом реплікації клітинної ДНК. Оскільки типи і форми вірусних нуклеїнових кислот дуже різноманітні (ДНК, РНК, двоспіральні й односпіральні, лінійні і кільцеві молекули), то і механізми їх реплікації теж різні. Віруси різних родів і родин мають різні способи передачі генетичної інформації і реплікації.
Синтез вірусних ДНК здійснюється за допомогою ДНК-полімераз, джерела яких можуть бути різні. Так, в адено- і герпесвірусів, що містять двоспіральні молекули ДНК, у складі віріонів немає ДНК-полімераз, тому на одній з ниток її синтезується іРНК і транслюється в рибосомах при синтезі ДНК-полімерази. Далі завдяки участі цього ферменту з нуклеотидів клітини синтезується і будується друга комплементарна нитка ДНК, у результаті чого утворюються нові двоспіральні молекули ДНК. Процес реплікації молекул ДНК продовжується доти, поки в клітині не нагромадиться кількість, необхідне для численного потомства вірусної частки, що проникнула в клітину.
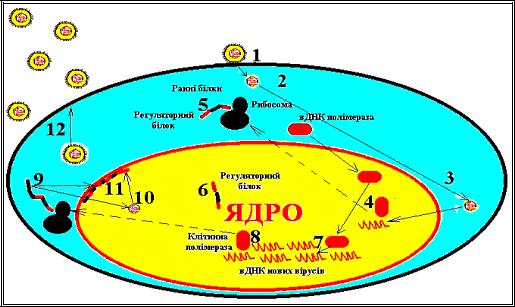
Реплікативний цикл ДНК – геномних вірусів. Після адсорбції (1)
вірус проникає в клітину шляхом злиття з мембраною (2), нуклеокапсид транспортується до ядерної оболонки (3), і вірусна ДНК ( вДНК) проникає в ядро, де починається її транскрипція (4). В результаті трансляції вірусного геному синтезуються ранні білки (5), включаючи регуляторні протеїни, вірусні полімерази і матричні білки. Вірусна полімераза проникає в геном клітини (6), де запускає синтез молекули ДНК дочірніх популяцій (7). Частина вірусної ДНК транскрибується клітинною РНК-полімеразою (8), що індукує синтез пізніх білків (9), необхідних для зборки дочірніх популяцій (10); останні залишають ядро, відбруньковуються від його мембрани (11) і включають її фрагменти в склад власної оболонки (12). Зрілі віріони транспортуються через цитоплазму і залишають клітину через комплекс Гольджі або після вибуху клітини.
37
Реплікація одноланцюгових вірусних ДНК.
Одноланцюгові вірусні ДНК у парвовірусів синтезуються по принципу комплементарності з проміжної реплікативної форми (ПРФ). У складі віріонів парвовірусів як структурний компонент є ДНК-залежна ДНК-полімераза. За допомогою цього ферменту за принципом комплементарності на вірусній односпіральній ДНК (плюс-нитці) утворюється одна комплементарна їй мінус-нитка. Надалі на цій двоспіральній структурі синтезуються дочірні плюс-нитки (матрицею для них є мінус-нитки).
Реплікація односпіральних вірусних РНК
(пікорнакорона- і арбовірусів). Реплікація вірусних РНК здійснюється без безпосередньої участі клітинних ДНК. Це підтверджується тим, що придушення синтезу ДНК і клітинної іРНК на матриці ДНК клітини не впливає на репродукцію багатьох РНК-містких вірусів.
Як відомо, одноланцюгові інфекційні РНК вірусів містять у собі всю генетичну інформацію, необхідну для синтезу вірусоспецифічних білків, і є, таким чином, інформаційними (іРНК) з матричними функціями. Після проникнення цих вірусів у клітину їх інфекційні РНК надходять безпосередньо в рибосоми і індукують синтез вірусних білків, у тому числі РНК-реплікази, які каталізують реплікацію самих вірусних РНК.
В односпіральних молекул РНК із кодуючими і матричними функціями (плюс-РНК) процес реплікації відбувається таким чином, що на вихідній вірусній молекулі РНК (на плюс-нитці) за участю ферменту РНКреплікази синтезується друга комплементарна мінус-нитка РНК, у результаті чого, утворюється двоспіральна реплікативна форма РНК. Інформація з плюс-РНК у рибосоми для синтезу вірусних білків передається безпосередньо плюс-РНК. Численні плюс-нитки вірусних інфекційних РНК увійдуть потім у нові частки вірусного потомства.
38
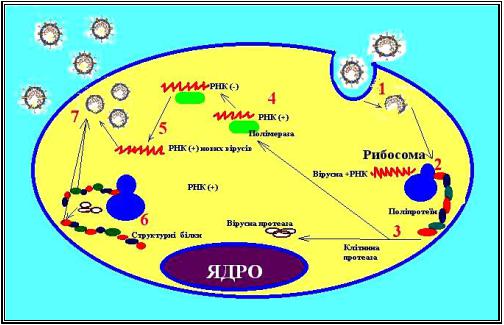
Реплікативний цикл +РНКгеномних вірусів.
Після адсорбції вірус проникає в клітину шляхом піноцитозу (1). Реплікативний цикл починається після звільнення вірусного геному в цитоплазмі, так як молекулярна симетрія вірусної РНК ( + РНК) аналогічна мРНК і вона може безпосередньо розпізнаватися і транслюватися рибосомами (2). Клітинні протеази трансформують утворений вірусний поліпротеїн (3) в РНК - залежну РНК - полімеразу, вірусну протеазу та різноманітні структурні білки. Полімераза копіює +РНК ланцюг в вигляді - РНК (4), яка виконує функцію матриці для синтезу молекули +РНК (5), які використовуються в синтезі вірусних білків (6), або білків що входять в склад генома нових популяцій вірусу (7).
В ортоміксовірусів (вірусів грипу А, В і С) на відміну від параміксовірусів геном представлений не однією молекулою нуклеїнової кислоти, а набором односпіральних РНК у виді восьми окремих, неоднакових по розміру субодиниць.
Розрізняють два різновиди РНК-полімерази: віріонну РНКтранскриптазу і РНК-репліказу. Перша відповідальна за переважний синтез мінус-ниток у ранній період інфекції, а друга— за переважний синтез плюсниток на більш пізніх етапах реплікації ортоміксовірусів.
Мінус-нитки утворюються на ранній стадії інфекції, і максимальний рівень їх синтезу передує появі плюс-ниток. Мінус-нитки РНК з'являються через 15 хвилин після зараження і незабаром виявляються у вірусоспецифічних полісомах. Мінус-нитки, синтезовані віріонною РНКтранскриптазою на ранній стадії інфекції, є іРНК. Установлено, що і деяка частина плюс-ниток виконує функції іРНК.
Надалі в зараженій клітині була виявлена ще одна форма вірусоспецифічної РНК. Молекули цієї форми містять двоспіральну «серцевину» і односпіральні "хвости". Ця форма одержала назву реплікативного попередника (РП) чи реплікативної проміжної форми (РПФ). Після депротеїнізації вірусу в клітині вивільнена РНК виконує функцію
39
іРНК, направляючи синтез вірус-специфічних білків, у тому числі РНКполімерази.
Синтез РНК може здійснюватися по одному з двох механізмів:
1)консервативному, при якому полінуклеотидні ланцюги, що входять до складу РПФ РНК, зберігаються (консервуються) і не переходять в односпіральну форму. Цей спосіб синтезу аналогічний способу синтезу односпіральних клітинних РНК на двоспіральній матриці ДНК;
2)утворення плюс-ниток може відбуватися асиметричним напівконсервативним шляхом, коли знову створена плюс-нитка витісняє раніше синтезовану плюс-нитку з РПФ РНК.
При використанні обох механізмів виникають проміжні структури типу РПФ. Кінцевим продуктом синтезу в обох випадках є односпіральна вірусна РНК, причому значна частина її бере участь у трансляції в складі реплікативного комплексу.
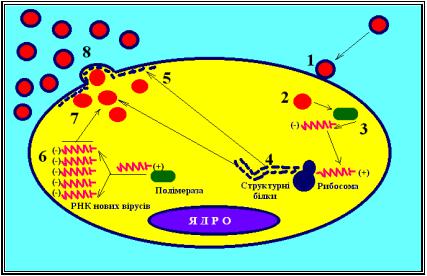
Реплікація односпіральних вірусних РНК із негативним геномом.
Реплікативний цикл – РНК геномних вірусів.
Є велика група односпіральних вірусів (рабдовіруси, пара-і ортоміксовіруси), що містять негативний (неінфекційний) геном. По складу, локалізації, функції й особливостям біосинтезу білків віруси цієї групи дуже подібні. У них як би єдина стратегія експресії генів. РНК їх (мінус-нитка) не транслюється в рибосомах. Функції іРНК у цих вірусів виконують плюснитки, комплементарні геномним. У складі вірусів з негативним геномом немає вірусоспецифічного ферменту типу РНК-реплікази, але міститься (крім вірусу сказу) РНК-залежна РНК-полімераза (РНК-транскриптаза). Остання синтезує на інфекційній мінус-нитці РНК комплементарні плюс-нитки іРНК у виді окремих фрагментів двох типів: перші мають полі (А) - послідовності на З’- кінці, надходять у рибосоми і служать матрицями для синтезу структурних білків і ферментів РНК-репліказ 1 і 2; другі не мають такої послідовності і служать матрицями для синтезу ниток віріонної РНК (мінусниток, що ввійдуть до складу нових часток вірусного потомства) за
40