

Конспект_лекцій_Заг.вірусологія_0
.pdfдопомогою цієї синтезованої РНК-реплікази 2.
Проникнення вірусу здійснюється після адсорбції і злиття з клітинною мембраною (1). Після роздягання (2) вірусна -РНК трансформується в плюс ланцюг РНК - залежною РНК-полімеразою, що входить в склад віріону (3), що призводить до утворення повних і коротких ланцюгів. Короткі +РНКланцюги зумовлюють синтез ферментів і білків для нових популяцій вірусів (4), серед останніх особливе значення має глікопротеїн оболонки (5), що вбудовується в клітинну стінку на етапах, які відбуваються до процесу відбруньковування. Повний ланцюг +РНК є матрицею для синтезу молекул - РНК нових популяцій вірусу (6). Нуклеокапсиди (утворюються із синтезованих білків) і -РНК прикріпляються до модифікованих ділянок клітинної стінки (7), відщеплюють фрагмент ліпідного шару (що завершує процес зборки вірусу) і відокремлюється шляхом брунькування (8).
Реплікація двоспіральних вірусних РНК.
Двоспіральні РНК містять реовіруси. Реовірус у складі віріону містить РНК-залежну РНК-полімеразу. Усі 10 фрагментів його генома транскрибуються зазначеною полімеразою. На одній з ниток (на мінус-нитці) синтезується односпіральна копія. Реплікація відбувається в такий спосіб. На мінус-нитці геномної двоспіральної РНК синтезується односпіральна плюснитка, що містить сайт зв'язування з рибосомою і сайт зв'язування з РНКзалежною РНК-полімеразою, який кодується вірусним геномом. Пул цих односпіральних (плюс-ниток) РНК служить спочатку матрицею для синтезу вірусних білків, у тому числі і згаданої полімерази. Після синтезу останньої вона зв'язується з цими ж плюс-нитками РНК і синтезує на їх матриці мінуснитку РНК. Цей процес обумовлений двома унікальними явищами:
-по-перше - одна і та ж молекула РНК служить матрицею для синтезу
ібілка і комплементарної РНК; - по-друге - структура РНК-залежної РНКполімерази реовірусів така, що по завершенні синтезу комплементарного ланцюга вона не відокремлюється від двоспіральної РНК. Як видно з викладеного вище, реплікація двоспіральних РНК має механізм, що у корені відмінний від механізму реплікації двоспіральних ДНК.
Вірусні двоспіральні РНК не можуть надходити в рибосоми безпосередньо. Укладена в них генетична інформація може передаватися в рибосоми опосередковано, тобто через односпіральні іРНК, оскільки в клітині немає вірусоспецифічного ферменту, що синтезував би односпіральну іРНК на матриці двоспіральній вірусної РНК, а також немає ферменту, що каталізував би реплікацію самої двоспіральної вірусної РНК.
Фермент, що синтезує односпіральну іРНК на матриці двоспіральної вірусної РНК, — РНКтранскриптаза — знаходиться в самих віріонах. Роль РНК-транскриптази зводиться до переписування генетичної інформації з двоспіральних вірусних РНК на односпіральні іРНК, тобто фермент РНКтранскриптаза виконує функцію, подібну функції ДНК-залежної РНКполімерази, що синтезує односпіральну іРНК на матриці двоспіральної ДНК клітини.
41
При передачі інформації з двоспіральних РНК у рибосоми, а також реплікації двоспіральних вірусних РНК РНК-транскриптаза синтезує на мінус-нитці двоспіральної РНК, як на матриці, комплементарну плюс-нитку і транскрибує на неї всю генетичну інформацію. Знову синтезована плюснитка РНК стає, таким чином, інформаційною вірусною РНК (іРНК). Остання надходить у рибосоми клітини, де індукує синтез вірусних білків, у тому числі РНК-реплікази. РНК-репліказа синтезує потім на плюс-нитці РНКкомплементарні мінус-нитки, і у результаті чого утворюються знову двоспіральні молекули РНК. Так забезпечується реплікація двоспіральних вірусних молекул РНК.
Реплікація ретровірусів
Уретровірусів для здійснення синтезу ДНК на матриці РНК необхідна наявність у заражених клітинах специфічного ферменту — РНК-залежної ДНК-полімерази (зворотної транскриптази), що міститься в складі вібріонів. Крім того, у складі віріонів виявлена ДНК-залежна ДНК-полімераза, яка здійснює синтез ДНК на матриці ДНК. Таким чином, в утворенні двоспіральної вірусоспецифічної ДНК беруть участь два різних ферменти. Зворотній транскриптазі належить, очевидно, провідна роль у процесах неопластичної трансформації клітин.
Ускладі віріонів, крім двох полімераз, є й інші ферменти, зв'язані з інтеграцією синтезованої ДНК із геномом клітини хазяїна. Сюди відносяться: ендонуклеаза, екзонуклеаза, полінуклеотидлігаза і рибонуклеаза Н.
Схема реплікативного циклу ретровірусів.
Утворення вірусоспецифічної ДНК починається на вірусній РНК, причому синтез здійснюється за допомогою ферменту РНК-залежної ДНК-
42
полімерази. У результаті утворюється гібридна двоспіральна молекула, одна нитка якої РНК, інша - ДНК. Далі, імовірно, відбувається вибіркове відщіплення нитки РНК-гібридної молекули за допомогою ферменту рибонуклеази Н.
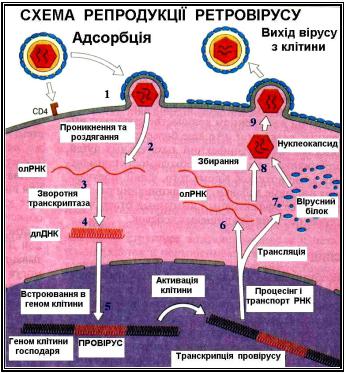
Інфекційна вірусна частинка (1) проникає в клітину шляхом злиття з клітинною мембраною після адсорбції на ній. Потім вірус роздягається (2), а зворотна транскриптаза, що входить в склад віріону індукує синтез - ДНК, яка використовує як матрицю молекулу РНК (3). +ДНК копіюється з нової синтезованої молекули - ДНК, в результаті чого утворюється подвійний ланцюг ДНК ( длДНК) (4). длДНК транспортується в ядро клітини, де клітина ДНК піддається сплайсингу з утворенням рекомбінантів з вірусною ДНК(5). Інтегрована молекула ДНК транскрибується клітинною ДНК - залежною РНК - полімеразою в +РНК , яка використовується в якості геномів нових популяцій (6), а також транслюється як мРНК для синтезу (через стадію утворення поліпротеїнів) структурних білків і ферментів (7). Деяка частина +РНК піддається сплайсингу з утворенням мРНК невеликого розміру, що кодує поверхневі, регуляторні та додаткові білки (8). Зрілі нові популяції вірусів вивільняються з клітини шляхом брунькування (9).
На односпіральній молекулі ДНК, що залишилася після руйнування РНК, синтезується комплементарна нитка ДНК, у результаті чого утворюється двоспіральна ДНК. Ця реакція здійснюється тією ж полімеразою, що бере участь в утворенні першої нитки ДНК. Потім двоспіральна ДНК вбудовується в клітинну хромосому, очевидно, за допомогою різних ферментів.
Вірусоспецифічна ДНК, убудована в клітинний геном, транскрибується з утворенням вірусної РНК, яка спочатку виконує функції іРНК, забезпечуючи синтез вірусних білків, а потім з'єднується з ними, формуючи нове покоління віріонів. На цьому цикл репродукції ретровірусів завершується.
ЛЕКЦІЯ 10. ЗБІРКА ВІРІОНІВ. ВИХІД ВІРУСНИХ ЧАСТОК ІЗ КЛІТИНИ
Загальні принципи збирання віріонів. Збірка РНК-вмісних вірусів.
Збірка ДНК-вмісних вірусів. Вихід вірусних часток із клітини
Синтез компонентів вірусних часток у клітині роз'єднаний і може протікати в різних структурах ядра і цитоплазми. Віруси, реплікація яких проходить у ядрах, умовно називають ядерними. В основному це ДНКвмістимі віруси: адено-, папова-, парвовіруси та віруси герпесу. Віруси, які реплікуються в цитоплазмі, називають цитоплазматичними. До них відносяться з ДНК-вмістимих вірус віспи і більшість РНК-вмістимих вірусів, за винятком ортоміксо- і ретровірусів. Однак цей поділ дуже відносний, тому
43
що в репродукції тих і інших вірусів є стадії, що протікають відповідно в цитоплазмі і ядрі. Усередині ядра і цитоплазми синтез вірусоспецифічних молекул також може бути роз'єднаний. Так, наприклад, синтез одних білків здійснюється на вільних полісомах, а інших - на полісомах, зв'язаних з мембранами. Вірусні нуклеїнові кислоти синтезуються в асоціації з клітинними структурами вдалині від полісом, що синтезують вірусні білки. При такому диз'юнктивному способі репродукції утворення вірусної частки можливо лише в тому випадку, якщо вірусні нуклеїнові кислоти і білки мають здатність при достатній концентрації дізнаватися один одного в різноманітті клітинних білків і нуклеїнових кислот і мимовільно з'єднуватися один з одним, тобто здатні до самозбирання.
В основі самозбирання лежить специфічне білок-нуклеїнове і білокбілкове впізнавання, що може відбуватися в результаті гідрофобних іонних і водневих зв'язків. Білок-нуклеїнове впізнавання обмежене невеликою ділянкою молекули нуклеїнової кислоти і визначається унікальними послідовностями нуклеотидів у некодуючій частині вірусного геному. З цього впізнавання ділянки генома вірусними капсидними білками починається процес збирання вірусної частки. Приєднання інших білкових молекул здійснюється за рахунок специфічних білок-білкових взаємодій чи неспецифічних білок-нуклеїнових взаємодій. Об'єднання білка з вірусними нуклеїновими кислотами в клітині відбувається спонтанно як чисто фізикохімічна реакція агрегації, що вимагає участі додаткових факторів (рН, іонної сили, іонів металів, осмосу і т.п.). Після того як концентрація вірусних РНК і білка досягає критичного рівня, у складно улаштованих вірусів принципи самозбирання забезпечує від початку до кінця морфогенез віріонів.
Дозрівання складних вірусів можна розділити на три етапи: 1) утворення внутрішньоклітинних нуклеокапсидів; 2) організація вірусної мембрани; 3) вихід зрілої вірусної частки з клітини за допомогою брунькування.
Загальні принципи збирання віріонів:
-У просто побудованих вірусів формуються провіріони, котрі в результаті модифікацій білків перетворюються у віріони. У складно побудованих вірусів спочатку формуються нуклеокапсиди або серцевини, з якими взаємодіють суперкапсидні білки.
-Збирання вірусів з суперкапсидною оболонкою ( за винятком покс - і гепаднавірусів) відбувається на клітинних мембранах: ядерній, якщо вірус реплікується в ядрі, або плазматичній, ендоплазматичної сітки чи комплексу Гольджі, якщо вірус реплікується в цитоплазмі. До цих мембран транспортується незалежно один від одного всі компоненти віріону.
-У деяких складно побудованих вірусів ( РНК - містких мінус - нитчастих ортоміксо-, параміксо- і рабдовірусів) існує спеціальний гідрофобний білок - матриксний, або мембранний. Він є медіатором складання віріонів: виконує посередницьку функцію між суперкапсидною оболонкою і нуклеокапсидом, утворюючи разом з ним серцевину.
44
- Складання нуклеотидів, серцевин, провіріонів і віріонів відбувається не у внутрішньоклітинній рідині, а в спеціальних структурах клітини - клітинних матриксах, віропластах. Ці структури локалізуються в ядрі або цитоплазмі інфікованих клітин і являються продуктами кооперативних процесів клітини й вірусу. Звичайно, це місця синтезу вірусних компонентів і складання віріонів потомства. У них виявляють різні клітинні структури - рибосоми ( полісоми) , мембрани, мікротрубочки, осміофільні волокна та ін.
- Складно побудовані віруси для побудови своїх віріонів використовують матеріал клітини-господаря, наприклад, ліпіди, вуглеводи, протеїнкінази, гістони, актин, тРНК(у ретровірусів), рибосоми (у аденовірусів). Клітинні елементи виконують певні функції в складі віріону.
Збірка РНК-вмісних вірусів.
Збірка простих вірусів полягає в асоціації вірусного генома з вірусними капсидними білками з утворенням нуклеокапсида.
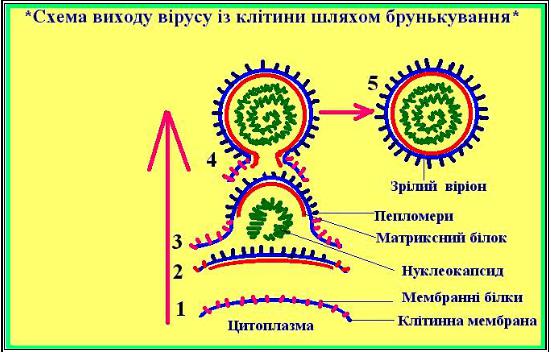
У складних вірусів процеси зборки нуклеокапсидів, серцевин і зрілих віріонів здебільшого роз'єднані. Нуклеокапсиди мігрують до місця зборки вірусних часток — плазматичної мембрани (чи мембран ендоплазматичної мережі) і упорядкування їх відбувається під ділянками мембран, із зовнішньої сторони яких вже убудовані вірусні суперкапсидні білки. Зборка полягає в тім, що ділянки, що містять глікопротеїди з пов'язаними з ними нуклеокапсидами, поступово випинаються через модифіковану клітинну мембрану. У результаті випинання утворюється «брунька», що містить нуклеокапсид і оболонку із суперкапсидними білками. «Брунька» відокремлюється від клітинної мембрани з утворенням вільної вірусної частки.
Брунькування може відбуватися через плазматичну мембрану клітини в зовнішнє середовище, як в ортоміксо-, параміксо-, рабдо- і альфавірусів, або через мембрани ендоплазматичної мережі у вакуолі, як в арена- і буньявірусів. В основі випинання бруньки через мембрану лежать звичайні клітинні процеси, спрямовані на відторгнення непридатного для клітини матеріалу і відновлення мембран. Ділянка майбутньої бруньки містить фіксований нуклеокапсид, асоційований з суперкапсидними білками; але рух мембранних ліпідів продовжується в силу їхньої плинності, ліпіди обволікають майбутню бруньку, і разом з ними із «бруньки» витісняються клітинні мембранні білки. У результаті цього руху відбувається вихід "бруньки" із клітинної мембрани. Механізм утворення "бруньки" пояснює, чому в складі вірусів, що брунькуються, не міститься клітинних мембранних білків.
Усі вірусні компоненти — нуклеокапсиди і суперкапсидні білки — прибувають до місця зборки незалежно один від одного. Першими до місця зборки прибувають суперкапсидні білки. Здебільшого цими білками є глікопротеїди, що синтезуються в полісомах. Через шорсткуваті, а потім гладкі мембрани в результаті злиття з ними везикул комплексу Гольджі ці білки транспортуються на зовнішню поверхню плазматичних мембран чи
45
залишаються в складі везикул. Включення глікопротеїдів у відповідні зони клітинних мембран призводить до модифікацій мембран. Нуклеокапсид впізнає ці ділянки і підходить до їх із внутрішньої сторони ліпідного шару. При зборці вірусів велику роль відіграє вірусний матриксний білок М, який є медіатором зборки. Включення М-білка в клітинні мембрани є сигналом для зборки вірусної частки: слідом за включенням негайно здійснюється зв'язування нуклеокапсидів з мембранами і брунькування вірусної частки. Тим самим М-білок має функцію фактора, що лімітує зборку.
Збірка ДНК-вмісних вірусів.
Узборці цих вірусів є деякі відмінності від зборки РНК-вмістимих вірусів. Як і у РНК-вмістимих вірусів зборка ДНК-вмістимих вірусів є багатоступінчастим процесом з утворенням проміжних форм, що відрізняються від зрілих віріонів по складу поліпептидів. Перший етап зборки полягає в асоціації ДНК із внутрішніми білками і формуванні серцевин чи нуклеокапсидів. При цьому ДНК з'єднується з попередньо сформованими "порожніми" капсидами.
Урезультаті зв'язування ДНК із капсидами з'являється новий клас проміжних форм, що називаються неповними формами. Крім неповних форм із різним вмістом ДНК, існує інша проміжна форма в морфогенезі — незрілі віріони. Зборка ядерних вірусів починається в ядрі, звичайно — в асоціації
зядерною мембраною. Проміжні форми герпесвірусу, що формуються в ядрі, брунькуються в перинуклеарний простір через внутрішню ядерну мембрану, і вірус здобуває таким шляхом оболонку, що є дериватом ядерної мембрани. Подальше добудування і дозрівання віріонів відбуваються в мембранах ендоплазматичної мережі й в апараті Гольджі, відкіля вірус у складі цитоплазматичних везикул транспортується на клітинну поверхню.
Вихід вірусних часток із клітини
Існує два способи виходу вірусного потомства з клітини: шляхом вибуху і шляхом брунькування. Вихід із клітини шляхом вибуху зв'язаний з деструкцією клітини, порушенням її цілісності, у результаті чого зрілі вірусні частки, що знаходилися усередині клітини, виявляються в навколишнім середовищі. Такий спосіб виходу з клітини належить вірусам, що не містять ліпопротеїдної оболонки (пікорна-, рео-, парво-, папова-, аденовіруси). Однак деякі із цих вірусів можуть транспортуватися на клітинну поверхню до загибелі клітини.
Вихід із клітини шляхом брунькування є характерним для вірусів, що містять ліпопротеїдну мембрану, які є дериватом клітинних мембран. При цьому способі клітина може тривалий час зберігати життєздатність і продукувати вірусне потомство, поки не відбудеться повне виснаження її ресурсів.
Вибухоподібний механізм звільнення віріонів пов'язаний з деструкцією (лізисом) зараженої клітини, в результаті чого вірусне потомство опиняється в позаклітинному просторі. Такий спосіб виходу властивий просто
46
організованим вірусам, які дозрівають і набувають інфекційної активності в середині клітини. Пригнічення метаболізму макромолекул (зниження синтезу клітинних нуклеїнових кислот і білків) і наступна деструкція зараженої клітини здійснюється структурними білками цих вірусів.
Вихід з клітини шляхом брунькування через плазматичну мембрану властивий більшості складно організованих вірусів. Даний механізм поєднує формування зрілих віріонів з одночасним звільненням їх з клітини. Це найбільш ефективний спосіб виходу, оскільки не залежить від деструкції заражених клітин. При цьому клітина може тривалий час зберігати життєздатність і продукувати вірусне потомство, доки не відбудеться повне виснаження її ресурсів. Включення вірусних глікопротеїдів у плазматичну мембрану надає клітині нової антигенної специфічності, і заражена клітина стає об'єктом дії імунних механізмів організму.
Схема виходу вірусу із клітини шляхом брунькування.
Деяким РНК - геномним вірусам, як вже зазначалося, властиве брунькування через мембрани ендоплазматичної сітки і комплексу Гольджі в середину цитоплазматичних вакуолей. Такі віруси як корона - і буньявіруси звільняються з зараженої клітини шляхом екзоцитозу: вакуолі зливаються з плазматичною мембраною, а віріони потомства опиняються в позаклітинному просторі. З допомогою екзоцитозу виходять з клітини ДНК - місткі герпесвіруси, котрі брунькуються через ядерну мембрану і транспортуються до поверхні клітини в мембранних везикулах. Звільнення з клітини механізмом екзоцитозу властиве також поксвірусам, які транспортуються в чохлах сформованих з мембран комплексу Гольджі.
У вірусу гепатиту В шляхом екзоцитозу виходить з клітини HBsAg, що призводить до антигенемії. Звільнення віріонів потомства зараженої клітини
47
може відбуватися на будь - якому етапі складання серцевини, оскільки в крові хворих виявляють не лише зрілі віріони, але й неповні віріони, що містять тільки вірусну ДНК або гібридні молекули ДНК/РНК.
Деякі віруси здатні виходити з клітини та вражати сусідні, обминаючи позаклітинний простір, що дозволяє їм уникати дії специфічних антитіл. Наприклад, герпесвіруси можуть проникати з однієї клітини в іншу по цистернах цитоплазматичного ретикулуму які з'єднують ядерну оболонку з плазматичною мембраною. Параміксовіруси при брунькуванні спричиняють злиття сусідніх клітин з утворенням гігантських багатоядерних клітин - симпластів ( синцитіїв).
Тривалість циклу репродукції вірусів - від моменту адсорбції до виходу з клітини - різна. Так у пікорнавірусів вірусів цей процес продовжується 5 - 10 годин, ортоміксовірусів 6 - 8 годин, реовірусів - 8 - 10 годин, аденовірусів 14 - 24 години, а у деяких їх представників - 32 -36 годин, герпесвірусів від 12 годин ( вірус хвороби Ауескі) до 70 годин і більше (цитомегаловірус людини). Швидкість розмноження вірусів колосальна. Наприклад, кількість інфекційного потомства одного віріону вірусу грипу через 8 годин досягає 103, а до кінця першої доби 1027. Урожай вірусу, тобто загальна кількість віріонів на одну заражену клітину, коливається в широких межах, зокрема, у пікорнавірусів - 25 - 100 тисяч, в аденовірусів - 10 000 - 1000000.
ЛЕКЦІЯ 11. ВЗАЄМОДІЯ ВІРУСУ З КЛІТИНОЮ
Основні типи взаємодії вірусу з клітиною Реакція клітини на вірусну інфекцію.
Основні типи взаємодії вірусу з клітиною
Існує два основних типи взаємодії вірусу і клітини. При першому типі вірусний геном функціонує в зараженій клітині більш-менш автономно. Репродукція його відбувається незалежно від репродукції клітинного генома. Віруси, що автономно розмножуються в клітині, відносяться до групи вірулентних. При такому характері взаємодії вірусу і. клітини утворюється нове покоління віріонів. У цьому випадку говорять про продуктивну взаємодію. Коли ж цикл репродукції переривається на якій-небудь проміжній стадії й інфекційного вірусного потомства не утворюється, така взаємодія вірусу і клітини називають абортивною. У тих випадках, коли симбіоз клітинного і вірусного геномів виявляється короткочасним і після утворення нового покоління вірусних часток заражена клітина (клітка-хазяїн) гине, таку реакцію на вірусну інфекцію називають літичною. Клітка гине під впливом вірусної інфекції від ряду незалежних одна від одної причин: 1) неспецифічного необоротного блокування клітинного генома на ранніх стадіях інфекції; 2) неспецифічних ушкоджень клітинного генома в процесі інфекції; 3) переключення метаболічних ресурсів клітини на вірусоспецифічні синтези; 4) специфічного лізису клітини при звільненні дозрілих віріонів у середовище; 5) порушення структури клітинних мембран
48
у результаті інтенсивного виходу вірусних часток у середовище. Явище, коли клітина, у якій автономно розмножується вірус, довгостроково зберігає свою життєздатність, одержало назву латенції.
Другий тип взаємодії вірусу і клітини властивий пухлинним вірусам, нуклеїнова кислота яких здатна вбудовуватися (інтегруватися) тим чи іншим способом у клітинну хромосому у формі провіруса, викликаючи трансформацію клітин. Границі між вірусами з автономною реплікацією геномів і інтеграційними вірусами дуже умовні, і один і той самий вірус у залежності від виду клітин може поводитися або як інфекційний, або як інтеграційний геном. Результатом такої взаємодії вірусу і клітини є зміна спадкоємних властивостей клітини. Цей тип взаємодії вірусу і клітини називають вірогенією, подібно лізогенії при взаємодії фагів з бактеріями. Віруси, здатні викликати вірогенію, відносять до групи помірних.
Реакція клітини на вірусну інфекцію.
Різні віруси можуть викликати найрізноманітніші клітинні реакції, пов'язаними зі змінами обміну і функції клітини. Клітинні реакції на вірусну інфекцію можуть бути чотирьох типів. До першої з них відносяться різноманітні патологічні зміни, що проявляються пригніченням синтетичних процесів, порушенням функціональної активності, пошкодженням структури самої клітини і її загибеллю. Такі зміни позначаються як цитопатична реакція на вірусну інфекцію, а здатність вірусів викликати таку реакцію — цитопатогенна дія. Другий тип клітинних реакцій полягає в синтезі закодованих у клітинному геноме білків (інтерферонів), що володіють антивірусною активністю. Третій тип реакції виявляється розмноженням вірусу без видимих патологічних змін клітин (латенція). І, нарешті, можливий четвертий тип, коли наявність вірусу в клітині супроводжується її проліферацією.
Зараження клітини, як правило, супроводжується пригніченням синтезу основних клітинних макромолекул — ДНК, РНК і білків. Однак пригнічення утворення цих трьох класів з'єднань настає не одночасно, а іноді вибірково. Найпоширеніше функціональне порушення зараженої клітини — це втрата здатності до поділу (пригнічення мітозу). По типі дії на мітотичну активність клітин віруси можна розділити на три групи: стимулюючі мітоз, інгібуючі мітоз і не впливають на мітотичну активність. Більшість міксовірусів придушує мітоз.
Віруси впливають і на хромосомний апарат клітини. При зараженні міксовірусами клітин виникають зміни хромосом, принаймні, трьох видів: окремі розриви, пульверизація хромосом, порушення правильної орієнтації клітинного веретена.
Відомі два способи утворення патологічних змін у клітині, зараженої вірусом. По першому з них цитопатогенну дія може викликати той чи інший структурний компонент віріону; при цьому зовсім не обов'язково, щоб геном вірусу проникнув у клітину. Такий спосіб дії вірусів називають цитопатогенною дією ззовні.
49
Якщо ж дія вірусу зв'язана з проникненням вірусного генома в уражену клітину й утворенням деяких продуктів, що і є першою причиною патології, то таку дію називають цитопатогенною дією зсередини.
Цитопатогенна дія ззовні описана для ряду вірусів тварин. Деякі віруси, що мають зовнішню ліпопротеїнову мембрану, на ранніх стадіях взаємодії з клітиною індукують утворення симпластів, чи полікаріоцитів, — злиття клітин у комплекси, що містять два чи кілька ядер. Симпластоутворення — результат злиття, при якому відбувається утворення багатоядерних клітин шляхом об’єднання цитоплазми клітин-сусідів. Виникаючі полікаріони можуть містити до 100 ядер. Для утворення полікаріонів потрібна досить велика множинність зараження (близько 1000 віріонів на клітину), відповідний сольовий склад середовища, зокрема обов'язкова присутність Са2+, і відповідна температура. Інтенсивність злиття залежить і від фізіологічного стану клітин. Описано два види злиття: ззовні і зсередини. Утворення симпластів відбувається в кілька стадій. Спочатку вірус викликає аглютинацію клітин, утворюючи як би місток між двома клітинними мембранами. Потім мембрани двох сусідніх клітин, розташованих поблизу віріонів, приходять у тісний контакт одна з одною. Далі (при наявності оптимальних умов) клітинні мембрани руйнуються в тім місці, де вони контактують з віріонами. У результаті утворюється канал між двома сусідніми клітинами. Поблизу утвореного отвору розірвані мембрани сусідніх клітин з'єднуються між собою, утворюючи цитоплазматичні містки між клітинами. Ці містки розширюються, і, нарешті, сусідні клітини цілком зливаються.
Під впливом вірусної інфекції in vitro можуть зливатися не тільки однакові, але і різні клітини того самого виду, а також клітини різних видів і навіть класів тварин. Утворені при цьому «гібридні» клітини одержали назву гетерокаріонів. Оскільки останні можуть бути життєздатними протягом тривалого часу, їх використовують для різноманітних цитологічних, біохімічних і генетичних досліджень.
Крім симпластів, віруси можуть індукувати у заражених клітинах утворення включень. Вірусні включення являють собою видимі під мікроскопом маси, що містять скупчення або вірусних часток (віруси групи віспи), або попередників вірусу (білків чи нуклеїнових кислот), або змінених клітинних компонентів. У залежності від складу ці маси можуть фарбуватися різними барвниками — кислими (оксифільні включення) чи основними (базофільні включення). Базофільні включення можуть являти собою місця зборки віріонів, а оксифільні відображають реакцію клітини на його вторгнення. Вірусні включення мають різноманітну, але характерну для даного вірусу локалізацію в клітині. Вони можуть бути важливою, а іноді єдиною ознакою вірусної інфекції в клітині.
Таким чином, патологічні порушення структури і функції інфікованої клітини можуть бути дуже різноманітні. У розвитку клітинних ушкоджень на пізніх стадіях інфекції відіграють роль не тільки вірусоспецифічні, але і клітинні білки — ферменти.
50