

Лекция 27
МЕХАНИЗМЫ ОБРАЗОВАНИЯ И ИСПОЛЬЗОВАНИЯ
АТР В ЖИВЫХ СИСТЕМАХ
Представления о механизмах сопряжения окисления и
фосфорилирования в дыхательной цепи. Хемиосмотическая теория
Митчелла. Электрохимический протонный градиент как форма
запасания энергии
Долгое время вопрос о механизме преобразования энергии, освобождающейся при переносе высокоэнергетических электронов по цепи окислительных ферментов, оставался неясным. Согласно хемиосмотической теории П. Митчелла (рис. 27.1), сопряжение тканевого дыхания с окислительным фосфорилированием обеспечивается внутренней митохондриальной мембраной, целостность которой обуславливает возникновение движущей силы синтеза АТР-протонного потенциала. В результате происходит перекачивание протонов из матрикса на цитоплазматическую поверхность и создается градиент рН. Движение протонов в обратном направлении (по каналу фактора Fo) ведет к активации АТР-синтазы (фактор F1) и синтезу АТР из АDP и фосфата. Транспорт АТР из матрикса в цитоплазму осуществляется переносчиком – транслоказой. Этот фермент катализирует перенос одной молекулы АТР из матрикса в обмен на одну молекулу АDP, переносимую в матрикс. Нарушение транспорта АDP или фосфата приводит к торможению синтеза АТP.

Рис.27.1. Английский биохимик Питер Деннис Митчелл (родился в Митчеме, в семье служащего Кристофера Гиббса Митчелла и Беатрис Дороти Митчелл, окончил Королевский колледж в Тонтоне и колледж Иисуса Кембриджского университета, в 1978 г. за разработку хемиосмотической теории удостоен Нобелевской премии)
Гипотеза П.Митчелла требует соблюдения ряда условий:
1) внутренняя митохондриальная мембрана должна быть интактна и непроницаема для протонов, направляющихся снаружи внутрь;
2) в результате активности дыхательной цепи ионы водорода поступают в нее изнутри, из матрикса, а освобождаются на наружной стороне мембраны;
3) движение ионов водорода, направленное изнутри наружу должно приводить к их накоплению, вследствие чего между двумя сторонами митохондриальной мембраны возникает градиент рН;
4) необходимы затраты энергии, которая поставляется при переносе электронов по электронтранспортной цепи;
5) синтез АТР поддерживать наличием электрохимического градиента.
Строение атp-синтазного комплекса. Механизм образования атp. Обратимость реакции, катализируемой атp-синтазой. Разобщение транспорта электронов и синтеза атp; действие 2,4-динитрофенола
В настоящее время протонные АТР-синтазы выделены практически из всех типов сопрягающих мембран: митохондрий, хлоропластов, хроматофоров. АТР-синтазный комплекс (Н+-АТРаза) – обратимый фермент, обладающий как АТР-синтазной, так и АТРазной активностью. Синтез АТР осуществляется за счет протонного градиента ΔμН+, а гидролиз АТР приводит к тому, что протонная АТРаза сопряженно генерирует трансмембранную разность электрохимического потенциала Н+. В Н+-АТРазе происходят процессы по общей схеме:
АТР-синтаза
ΔμН+ ↔ АТР
АТРаза
АТР-синтазный комплекс (F0F1-АТРаза) состоит из растворимой АТР-синтазы (фактор F1) и мембранных компонентов (комплекс F0) (Рис.27.2).
Сопрягающий фактор АТР-синтазы (F1 для митохондрий и CF1 для хлоропластов) представляет собой полифункциональный белок, имеющий сложную четвертичную структуру. Он построен из трех типов крупных субъединиц – α, β, γ с молекулярной массой 30000-60000 Да и двух типов минорных субъединиц δ, ε с молекулярной массой 11000-20000 Да. Стехиометрия комплекса – 3α3βγδε. Разложение его на субъединицы ведет к потере ферментативной активности. «Шляпка» высотой 80Å и шириной 100Å грибовидного выроста АТР-синтазы соответствует фактору F1, частично погруженному в мембрану, в основании которого находится гидрофобный белок комплекса F0, который включает 3 типа полипептидов (a,b,c) с молекулярными массами от 6500 до 30000 кДа и обеспечивает связывание фактора F1 с мембраной и перенос протонов при работе фермента.

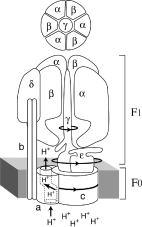
Рис.27.2. Общая топография АТР-синтазного комплекса
F1 – сопрягающий фактор, Fo – протонный канал (пояснения в тексте)
Комплекс F1 может поворачиваться вокруг оси, совпадающей по направлению с субъединицей γ. При повороте на 120º каждая из α и β-субъединиц перемещается на место другой такой же, а при повороте на 60º субъединицы меняются местами. В силу некоторой асимметричности вращение F1-комплекса переводит его β-субъединицы при каждом повороте в новое положение, где они попадают в другое микроокружение. Это и приводит к изменению состояния и конформационной перестройке активных центров при вращении F1. Каждый из трех активных центров в результате вращения F1-комплекса может поочередно находиться в одном из трех конформационных состояний, которые различаются по степени сродства молекул АТР, АDP и Pi к каталитическому центру. В состоянии 1 центр β-субъединицы открыт и в нем связываются молекулы АDP и Pн, которые сравнительно слабо удерживаются центром.
Вращение и конформационные перестройки комплекса F1 переводят этот центр в состояние 2, где непосредственно происходит синтез АТP. Здесь АDP и Pн прочно фиксируются в каталитическом центре, находясь в активной конфигурации, необходимой для образования ковалентной связи между фосфатными группами АDP и Pн. Поэтому на этой стадии не требуется притока энергии извне. В состоянии 2 в каталитическом центре самопроизвольно идет образование и разрыв ковалентной связи АDP-P, поскольку константа равновесия АТP↔АDP+Pi близка к единице.
На следующем этапе центр переходит в состояние 3, где за счет энергозависимой структурной перестройки происходит ослабление прочной связи молекулы АТP с центром и выход ее наружу. На освободившееся место из раствора приходят новые молекулы АDP и Pн. Перестройки носят кооперативный характер и затрагивают состояние всех трех каталитических центров β-субъединиц. Поскольку весь цикл включает 3 этапа, а в Н+-АТР-синтазе имеется 3 субъединицы, то после каждого структурного перехода в растворе появляется новая молекула АТР.
F0 формирует канал, по которому протоны поступают к активному центру АТРазы. Проводимость протонов носит специфический характер и подавляется антибиотиком олигомицином и ДЦКД (N,N′-дициклогексилкарбодиимид), ингибиторами Н+-АТР-синтазы. Особым образом организованный канал обеспечивает прохождение протона через всю мембрану из водной фазы в гидрофобную область мембраны, а затем из нее в воду по другую сторону от липопротеинового барьера. Основную роль в переносе играет ДЦКД-связывающий протеолипид. Предполагают, что он располагается поперек мембраны, так что полярная часть оказывается на внешней поверхности мембраны и служит входом в канал. Наиболее вероятным механизмом переноса протона представляется эстафетная передача по протон-донорным и протон-акцепторным группам аминокислот, включая остатки аргинина, тирозина, глутамина.
Комплекс F0 ‒ вращающийся ансамбль субъединиц с 9-12 остановками, позволяющими прерывать поток протонов. Временное связывание протонов может осуществляться остатком аспарагиновой кислоты на каждой субъединице. Каждый поворот γ-субъединицы внутри комплекса происходит в соответствии с тем, заполнено ли гидролитическое место АТР или АDP и Pн. Эти конформационные изменения (механические изменения структуры) обеспечивает перенос протонов водорода на каждый третий этап цикла. Связывание, гидролиз и освобождение АТP и ADP в F0-F1-АТP-синтазе зависит от величин и геометрии расположения заряда в активном состоянии. Пока еще не ясна природа и расположение заряженных групп, ответственных за непосредственное вращение молекулярного мотора Н-АТP-синтазы.