

Иммунологиялық жад, иммунологиялық толеранттылық
Иммунологиялықжадантигеннің бастапқы енуі (бастапқы иммундық жауап) кезінде пайда болатын Т- және В- жад жасушаларының болуына негізделген. Жад жасушалары өзіне тән антигеннің әсерінен тез пролиферацияланады: эффектор- жасушалардың көп популяциясы пайда болады, антиденелердің және цитокиндердің түзілуі жоғарылайды. Жад жасушаларының әсерінен қайтадан енген антигендер (екіншіреттегі иммундық жауап кезінде) тез және нәтижелі түрде жойылып отырады. Екінші реттегі иммундық жауап кезінде IgG-дің пайда болу жылдамдығы, саны және аффинділігі біршама артады (23-сурет). Иммунологиялық жад кейбір жұқпалар (жел шешек, қызамық және т.б.) кезінде жылдар бойы және өмір бойы сақталуы мүмкін.

23-сурет. Бірінші және екінші иммундық
жауап кезіндегі антидене түзілуі.
Иммунологиялық толеранттылық – организмде лимфоциттерге қолайлы антигендер (толерогендер) болған жағдайда иммундық жауаптың болмауы. Көбінесе ерігіш антигендер толерогенді болып келеді, өйткені антигентаныстырушы жасушаларда иммунды жауапқа сәйкес костимулдаушы молекулалардың экспрессиясын тудырылмайды.
Табиғи иммунологиялық толеранттылық – лимфоциттердің аутореактивті клондарының теріс сұрыпталуымен сипатталады. Организмнің өзіне меншікті антигендеріне толеранттылығы лимфоциттердің аутореактивті клондарының онтогенез процессікезіндежойылып отыру нәтижесінде дамиды. Т-лимфоциттер тимуста теріс сұрыпталуға ұшырайды, ал көптеген В-лимфоциттер сүйек кемігінде ұшырайды. “Тосқауыл сыртындағы ағзалар” саналатын антигендер қалыпты жағдайда аутоиммунды жауап тудырмайды, өйткені иммундық жүйе жасушаларымен қатынаспайды; жарақаттану, ұзақ жұқпалы аурулар кезінде бұл антигендер қанға түсіп “тосқауыл сыртындағы ағзалар” антигендеріне қарсы иммундық жауап тудырады.
Жасанды иммунологиялық толеранттылық –ұрыққа немесе нәресте туылғаннан кейін бірден (яғни, иммунологиялық жетілмеген кезеңде) бөгде антигендерді енгізген кезде; сәулелендіру цитоуытты агенттерді және т.б. енгізу нәтижесінде иммунды жүйенің басылуы немесе бұзылуы кезінде туындауы мүмкін.
Иммунологиялық толеранттылық келесі бағыттар бойынша дамиды: антигенді өзінің рецепторларымен байланыстырған лимфоцит клонының делециясы және (белсену орнына) апоптоз сигналы нәтижесінде өлуі; антигенді өзінің Т- және В-жасушалық рецепторларымен байланыстырған лимфоцитердің белсенуінің жоқтығынан лимфоциттер клонының анергиясы. Егер антигенді таныстыру кезінде антигентаныстырушы жасушаның бетінде костимулдаушы В7 (СД80 және СД86 ) молекуласы экспрессияланбаса Т-лимфоцит антигенге жауап бермейді.
Аллергия: жсжт, жсбт
Аллергия (грек тілінен allos − бөтен) − иммундық жүйенің талапқа сай емес реакциясы нәтижесінде антигендерге өзіндік жоғары сезімталдық болып табылады. Аллергия жоғарысезімталдықтың жедел және баяу түрінде көруі мүмкін.
-Жоғарысезімталдықтың жедел типі (ЖСЖТ) аллергендерге қарсы антиденелердің IgE,IgG,IgM – қалыптасуы (3-кесте). Аллерген әсер еткеннен кейін бірнеше минут немесе сағаттан кейін дамиды: қан тамырлары кеңейіп, олардың өткізгіштік қасиеті жоғарылайды, қышыну, бөртпе, бронхоспазм, ісінулер пайда болады. ГСЖТ-ның кешіктірілген фазасы эозинофилдер мен нейтрофильдердің өнімдерінің әсерімен толықтырылады.
-ЖСЖТ-не аллергиялық реакциялардың (Джелл мен Кумбс бойынша) І, ІІ, ІІІ типтері жатады: І типі – анафилаксиялық, негізінен IgE –дің әсерімен сипатталады; ІІ типі – цитоуытты, IgG, IgM әсерімен сипатталады; ІІІ типі – иммунокешендік, IgG, IgM−нің антигендермен иммундық кешен түзуінен дамиды. Сонымен қатар антирецепторлы реакция типі де қарастырылған.
-Жоғарысезімталдықтың баяу типі аллергияның (Джелл мен Кумбс бойынша) ІV типіне жатады. Бұл түрі жасушалық иммунитетті белсендіретін антигеннің (аллергеннің) макрофагтармен және Тһ1-лимфоциттермен әсерлесуі арқылы жүреді. Негізінен, аллерген енгеннен кейін 1−3 тәуліктен кейін дамиды: оның Т-лимфоциттермен және макрофагтармен инфильтраттар пайда болуы нәтижесінде тіннің тығыздалуы мен қабынуы дамиды.
Жоғарысезімталдық реакцияларының негізгі типтері.
І типі − анафилаксиялық. Антигенмен бірінші байланысқан кезде IgE пайда болады, олар Ғс-фрагменті арқылы мес жасушалар мен базофильдерге жабысады (24-сурет). Екінші рет енгізілген антиген жасуша бетіндегі IgE-мен айқаса байланысып, жасушаның дегрануляциясын тудырып, гистамин және басқа да аллергиялық медиаторларды бөледі.

24-сурет. Анафилакиялық типі
ІІ типі − цитоуытты. Жасушада орналасқан антиген IgG, IgM класты антиденелерімен танылады. “Жасуша-антиген-антидене” түрінде өзара әрекеттескенде комплементтің белсендірілуі жүреді және жасушаның бұзылысы үш бағыт (25-сурет): комплемент-тәуелді цитолиз (А); фагоцитоз (Б); антидене-тәуелді жасушалық цитоуыттылық әсер (В)бойынша жүреді.

25-сурет. Цитоуытты типі.
ІІІ типі − иммунокешенді. IgG, IgM кластыантиденелер еритін антигендермен байланысып, комплементті белсендіретін иммундық кешендер қалыптастырады.
Антигендердің артық болуынан немесе комплементттің жеткіліксіздігінен иммундық кешендер тамыр қабырғаларына, базальдық мембраналарға, яғни, Ғс рецептоплары бар құрылымдарға жиналады (26-сурет).
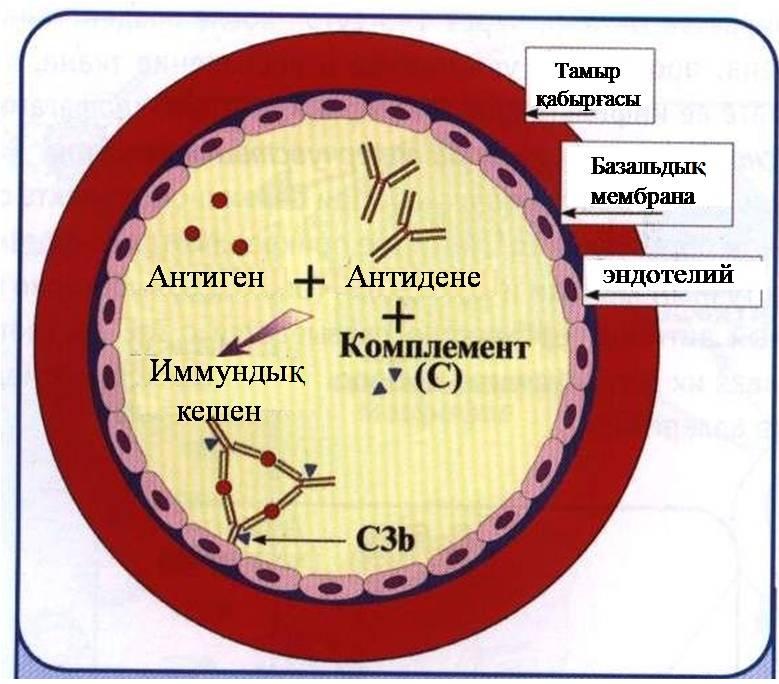
26-сурет. Иммунокешендік типі
ІV типі − жоғарысезімталдықтың баяу типі. Бұл түрі жасушалық иммунитетті белсендіретін антигеннің макрофагтармен және Тһ1-лимфоциттермен өзара әрекеттесуі арқылы жүреді (27-сурет).

27-сурет. Гиперсезімталдықтың баяу типі. ГСБТ
Жоғарысезімталдықтың І типі – анафилаксиялық, аллергеннің бірінші түсуі плазмоциттерденIgЕ, IgG4 өнімінің бөлінуін тудырады
IgЕ -антиденелерінің өндірілуін Тһ2 бөлетін ИЛ-4 және ИЛ-10 ынталандырады, ал Тһ1 бөлетін – гамма интерферон және ИЛ-2 тежейді. Синтезделген IgЕ Ғс-фрагменттері арқылы қан базофилдерінің және сілемейлі қабықтардағы, дәнекер тініндегі мес жасушаларының Ғс-рецепторына (ҒсεR1) бекінеді. Аллергеннің екінші рет түсуі кезінде мес жасушалар мен базофилдерде жасушалардың дегрануляциясын тудыратынIgЕ−дің аллергенмен (ҒсεR1-дің антигенмен айқасуы) кешеніқалыптасады (28-сурет).
Гранулалардан тіндерге биологиялық белсенді медиаторлар бөлінеді: вазоактивті аминдер (гистамин), протеогликандар (гепарин), липидтік алмасудың өнімдері (лейкотриендер, простогландиндер және тромбоциттерді белсендіруші факторлар), ферменттер (триптаза, химаза, карбоксипептидаза, Gкатепсині) және цитокиндер (Ил-3, ИЛ-4, ИЛ-5, ИЛ-6,ИЛ-13, ГМ-КСФ, ФНО-α).
Хемотаксистік факторлар нейтрофилдерді, эозинофилдерді және макрофагтарды тартады. Эозинофилді хемотаксистік факторлар (ЭХФ) эозинофилдерді тартады, олар ферменттерді, катионды нәруыздарды, лейкотриендерді және эпителийді зақымдаушы негізгі нәруыздеарды бөледі.
Аталған компоненттер тегіс салалы бұлшықеттердің жиырылуын, жүрек жұмысының әлсіздігін, коллапс дамуын, қан тамырлар өткізгіштігінің артуын, ісінулердің, қышынулардың пайда болуын және т.б. тудырады.
Жоғарысезімталдықтың І типінің клиникалық көріністері.
Жоғарысезімталдықтың І типінің клиникалық көріністері атопия кезінде жүруі мүмкін. Атопия – жоғарысезімталдықтың жедел түрінің дамуы аллергенге IgЕ-антиденелерінің шамадан артық түзілуімен, мес жасушасының бетіндегі осы антиденелерге Ғс-рецепторларының санының артуымен, мес жасушаларының таралуының ерекшеліктерімен және тіндік барьерлердің өткізгіштігінің жоғарылауымен сипатталатын тұқымқуалаушы бейімділік.
Анафилаксиялық шок – жедел дамып, коллапстың, ісінулердің, тегіс салалы бұлшықеттердің спазмы дамуымен жүреді; көп жағдайда өліммен аяқталады. Есекжем – қантамырларының өткізгіштігі артады, тері қызарады, көпіршіктер, қышыну пайда болады. Ангиоэдема – теріасты және субмукозды тіндердің ісінуі; көп жағдайда есекжеммен бірлесіп жүреді. Аллергиялық ринитнемесе аллергоконъюктивит – ісіну дамып, сілемейлі қабық тітіркенеді.Поллиноз – өсімдіктердің шаң −тозаңдарына аллергия. Сілемейлі қабықтардың вазомоторлық көріністерімен (риниттерде, конъюктивиттерде) және бронхоспазммен жүреді. Бронх демікпесі – қабыну, бронхоспазм дамып, бронхтағы сөлдің секрециясы жоғарылайды. Тағамдық аллергия – жүрек айну, диарея, қышыну, бөртпе, анафилаксия.
Прауснитц-Кюстнер реакциясы. Жоғарысезімталдықтың І типін антиденелердіңкөмегімен пассивті тасымалдауға болады. Ең алғашнауқастың қан сарысуымен аллергияның иммунореагентін пассивті тасымалдауды неміс бактериологы Прауснитц көрсеткен. Ол өзінің терісіне балыққа аллергиясы бар Кюстнердің (неміс гинекологы) қан сарысуын енгізген. Терінің аллерген енгізілген жеріне сәйкес аллергенді енгізген кезде ісіну, көпіршіктердің пайда болуы байқалған. Бұл сынамаПрауснитц-Кюстнер реакциясы деп аталды: науқас адамнан сау адамға енгізілген аллергенге IgЕ-антиденелер мес жасушалардабекітіледі де, аллергенді қайта енгізген кезде осы мес жасушаларымен оның байланысуы жүріп, гистамин және басқа белсенді субстанциялар бөлінеді (29-сурет).
3-кесте.Аллерген түрлері−аллергиялық реакция тудыратын төмен молекулалы нәруыздар немесе гаптендер
|
Ингаляциялық |
а) өсімдік тектес, мысалы өсімдік тозаңдары ринит, каньюктивит және бронхо спазм түрінде поллиноз тудырады. |
|
б)жануар тектес (эпидермальды антигендер, кене антигендері және т.б) | |
|
в) тұрмыстық аллергендер (шаң және т.б) | |
|
Тағамдық |
жұмыртқа, сүт, сыр, балық, ет, шоколад, жаңғақ, жемістер, көкшөп, тәттілер, жидектер, тағамдық қоспалар |
|
Дәрілік |
Антибиотиктер, сульфаниламидтер, гормондар (инсулин АКТГ, ТСГ), сарысулар, витаминдер (тиамин және т.б) ферменттер және т.б |
|
Инфекциялық |
Бактериялар, саңырауқұлақтар, қарапайымдылар антигені |
|
Өндірістік |
Полимер, пестицид, металлдар және т.б |
Лабораториялық диагностика. Науқас қанындаболжамды аллергендерге жалпы IgЕ, IgЕ- және IgG антиденелерін, гистамин, триптаза, интерлейкиндердің (ИЛ-5, ИЛ-4) деңгейін анықтау. Поллинозбен ауыратын науқастарда асқыну кезінде мұрын қуысынан алынған жағындыда эозинофилдердің саны 10%-дан 100 %-ға дейін жоғарылайды (қалыпты жағдайда − 2%-дан аспайды). Қанда да эозинофилия болуы мүмкін. Атопиялық аллергендермен тері сынамалары (30-сурет): шаң-тозаң, тұрмыстық, тағамдық, эпидермалды т.б. Провокациялық назальды, ингаляциялық және т.б. сынамалар.

28-сурет.Жоғарысезімталдықтың Ітипі

29-сурет.Аллергиясыбарнауқастанқансарысуымен
аллергияны тасымалдау (Праусниттің-Кюстнер реакциясы

30-сурет.Скарификациялық терілік сынама. Аллергенді енгізген соң (15−20мин.соң) реакцияны бағалайды: реакция оң болса−күлдіреуік (2 ден 10мм-ге дейін және одан жоғары) және қызару пайда болады. Теріс нәтижеде−күлдіреуік, айқын гиперемия болмайды (нәтижесін аллерген ерітіндісін және гистаминге реакциямен салыстырады)
Жоғарысезімталдықтың ІІ типі − цитоуытты. Жасуша мембранасына бекінген эндогенді антигендер немесе экзоненді химиялық заттар, дәрілік препараттар (гаптендер) жоғарысезімталдықтың ІІ типіне әкелуі мүмкін. ОлIgM немесе IgG кластыантиденелерінің және комплементтің (комплемент -тәуелді цитолиз) қатысуымен жүруі мүмкін. Сондай-ақ фагоциттер және К-жасушалары да антиденеге тәуелді жасушалық цитоуыттылық (АТЖЦ) түрінде қатысуы мүмкін. Реакция уақыты −минуттар немесе сағаттар.
Жоғарысезімталдықтың ІІ типіне антирецепторлық антиденелер негізі болып табылатын антирецепторлық реакциялар (жоғарысезімталдықтың V типі деп аталатын) ұқсас.Мысалы, гормондарға арналған рецепторларға қарсы антиденелер.
Цитоуыттытипінің түрлеріне: цитолиз,фагоцитоз және антиде−тәуелді жасушалық цитотоксикалық әсержатады.
- Комплементпен жанамаласқан цитолиз. Антиденелер жасуша бетіндегі антигендерге бекінеді.Кейінненантиденелердің Ғс-фрагментіне комплемент (С) қосылып, анафилатоксиндер (С3а, С5а) және C5−9 компоненттерінен тұратын мембранашабуылдаушыкешенніңтүзілуімен классикалық жолыбелсендіріледі (6-суретті қараңыз). Комплементке тәуелді цитолиз жүреді (31-сурет).
- Фагоцитоз. Фагоциттер антиденелермен және комплементпен (С3b) опсанизацияланған антигені бар нысана жасушаларды жұтуғажәне (немесе) бұзуға қабілетті (31-сурет).
- Антиденеге-тәуелді жасушалық цитоуыттылық әсер. Антиденелермен опсонизацияланған нысана-жасушалардың ТЖ-жасушаларымен лизисі. ТЖ-жасушалары нысана-жасушалардың антигендерімен байланысқан иимуноглобулиндердің Ғс-фрагментіне қосылады (32-сурет).
Клиникалық көріністері. Жоғарысезімталдықтың ІІ типі бойыншаөз тіндерінің антигендеріне қарсыаутоантиденелердің пайдаболумен жүретін кейбір аутоиммундық аурулар дамиды: қатерлі миастения (33-сурет), аутоиммундық гемолитикалық анемия, кәдуілгі күлдіреуік, Гудпасчер синдромы, аутоиммундық гипертиреоидизм, инсулинге тәуелді ІІтипті диабет.

31-сурет.Комплементке тәуелді цитолиз және фагоцитоз
Аутоиммунды гемолитикалық анемияны эритроциттердің Rh антигеніне қарсы антидене тудырады;эритроциттер комплемент және фагоцитоздың белсенуі нәтижесінде ыдырайды. Дәрілік-индукцияланған гемолиздік анемия,гранулоцитопения және тромбоцитопения дәрі-гаптенге қарсыантиденелердің пайда болумен және осы антигені бар жасушалардың цитолизімен жүреді.

32-сурет.Антиденегетәуелдіжасушалықцитоуытты әсер.

33-сурет. Қатерлі миастения кезіндегі антирецепторлық антиденелер.
Кәдуілгі күлдіреуік (теріде және сілемейлі қабықтардағы көпіршіктер түрінде) жасушааралық адгезия молекулаларына қарсыаутоантиденелер тудырады. Инсулинге тәуелді ІІ типті диабет инсулинге арналған рецепторларды тежейтін аутоантиденелермен тудырылып, гипергликемиямен және кетоацидозбен бірлесе жүреді. Гудпасчер синдромы бүйрек шумақшаларының капиллярларының базальды мембранасына қарсы аутоантиденелер түзіліп, өкпедегі қанқұйылулармен қоса жүретін нефрит дамиды. Айқын әлсіздікпен жүретін қатерлі миастениякезінде бұлшықет жасушаларының бетіндегі ацетилхолин рецепторларына қарсы антиденелер (аутоантиденелер) пайда болады. Антиденелер ацетилхолиннің рецепторлармен байланысуын тежейді де бұлшықет әлсіздігі туындайды. Басқа аутоантиденелер, керісінше, тежеудің орнына белсендіруші әсер етеді. Мысалы, аутоиммундық гипертиреоидизм (Грейвс синдромы) кезінде ТТГ-ға (тиреотропты гормонға) арналған рецепторларға қарсы антиденелер ТТГ-ның әсерін айнытпай қайталап, қалқанша безінің функциясын белсендіреді.
Лабораториялық диагностика. Айналымдағы тіндергеқарсы антиденелерді анықтау, сонымен қатар зақымдалған жерде (биопсия) антиденелер мен комплементті иммунофлюоресценция реакциясы арқылы анықтау.
Жоғарысезімталдықтың ІІІ типі − иммунокешендік. IgG, сирек жағдайда − IgМ қатысуымен жүретін ерігіш иммундық комплекстердің (антиген-антидене-комплемент) түзілуімен пайда болады (34-сурет).
Жоғарысезімталдықтың ІІІ типінің біріншілікті компоненттері ерігіш иммундық комплекстердің антиген-антидене және комплемент (анафилотоксиндер С4а, С3а, С5а) болып табылады. Антигендердің артық болуынан немесе комплемент жеткіліксіздігінен иммундық кешендер қантамырлар қабырғасына, базальды мембраналарға, яғни, Ғс-рецепторлары бар құрылымдарға тұнады. Зақымдалу тромбоциттермен, нейтрофилдермен, иммундық кешендермен, комплементпен сипатталады. Қабынуалды цитокиндері қосылады, соның ішінде ІНФ-α және хемокиндер. Кеш кезеңінде үрдіске макрофагтар қосылады.
Реакция жалпы болуы мүмкін (мысалы, сарысулық ауру) немесе жеке мүшелерді, тіндерді, теріні де (мысалы, жүйелі қызыл жегі, Артюс реакциясы), бүйректі (мысалы, жегілік нефрит), өкпені (мысалы, аспергиллез) немесе басқа мүшелерді қамтуы мүмкін. Бұл реакция көптеген микроорганизмдермен шақырылуы мүмкін. Ол Артюс реакциясындағы сияқты антиген экспозициясынан кейін 3−10 сағаттан кейін дамиды. Антиген экзогенді (созылмалы бактериалдық, вирустық, саңырауқұлақты немесе протозойлық жұқпалар) немесе жүйелі қызыл жегі кезіндегі сияқты эндогенді болуы мүмкін.
Сарысулық ауру антигеннің көп мөлшерін енгізгенде, мысалы, жылқының сіреспеге қарсы сарысуын енгізгенде, дамиды. 6−7 күннен кейін қанда жылқының нәруыздарына антиденелер пайда болып, сәйкес антигенмен әрекеттесіп, қантамырлардыңқабырғасына және тіндерге тұнатын иммундық комплекстер түзеді.Жүйелі васкулиттер, артриттер (кешендердің буындарға тұнуы), нефриттер (кешендердің бүйректерде тұнуы) дамиды.
Артюс реакциясы антигенді қайтадан теріішіне енгізген кезде дамиды, жергілікті бұрын жиналған антиденелермен иммундық комплекстер түзіледі. Ісінумен, геморрагиялық қабынумен және некрозбен көрініс береді.
Лабораториялық диагностика. Иммунофлюоресценция реакциясы арқылы иммуноглобулиндердің және комплементтің тұнуын анықтау үшін тіндердің биоптатын зерттейді. Қан полиэтиленгликолімен тұндырылған иммундық кешендердедегі IgG-ді анықтайды.

Сурет−34. Жоғарғысезімталдықтың ІІІ түрінің даму сатысы. (қантамыр қабатында иммундық кешендердің жиналуы)
4-кесте. Жоғарысезімталдықтың ІV типінің үш түрін ажыратады
|
ГСБТ түрлері |
Реакция уақыты |
Гистологиясы |
Клиникасы | |
|
Жанаспалы |
48−72 сағат |
Лимфоциттер, кешірек −макрофагтар |
Экзема. Ісінулер | |
|
Туберкулиндік |
48−72 сағат |
Лимфоциттер, моноциттер, макрофагтар |
Жергілікті индурация | |
|
Гранулематоздық |
21−28 тәулік |
Макрофагтар, эпителий жасушалары, гигантты жасушалар. Фиброз |
Теріде, өкпеде және т.б. тығыздалулар | |
|
Дәрілік заттар, косметикалық препараттар, төмен молекулалы заттар (гаптендер) тіндердің нәруыздарымен қосылып, жанаспалы ЖСБТ-нің дамуына әкелетін кешендік антигендерді тудырады. Жұқпалы аурулар (бруцеллез, туляремия, туберкулез, лепра, токсоплазмоз, көптеген микоздар және т.б.) ЖСБТ дамуымен қатарласып жүреді, сондықтан диагностика кезінде қоздырғыштардың антигендерімен терілік-аллергиялық сынамалар қолданылады (35-сурет): туберкулиндермен, лепроминмен, бруцеллинмен, туляринмен, токсоплазминмен және т.б. |
35-сурет.Теріге жасалған сынама. | |||

ІV типі − жоғарысезімталдықтың баяу типі (ЖСБТ). (4-кесте). Жасушалық иммунитеттіңбелсенуіне жауап беретін макрофагтар мен Тһ1-лимфоциттерінің қатысуымен жүреді (27-сурет).
ЖСБТ негізінен аллерген әсер еткеннен кейін 1−3 тәулік ішінде дамиды: тіннің Т-лимфоциттермен және макрофагтармен инфильтрациясы нәтижесінде тіннің тығыздалуы мен қабынуы дамиды.
Лабораториялық диагностика. Тері сынамаларын қоюмен сипатталады (аппликациялық немесе path-сынамалары), теріні гистологиялық зерттеу.
Цитокидер
Цитокиндер – жасушааралық қарым-қатынасты қамтамасыз ететін иммундық жүйенің белсендірілген жасушалары бөлетін нәруыздар. Оларға интерферондар (ИНФ), интерлейкиндер (ИЛ), ісік некроздаушы факторлар (ІНФ), колониястимулдаушы факторлар (КСФ), өсу факторлары, хемокиндер жатады.

36-сурет. Иммундық жауаптың аутокринді-паракринді реттелуі.
Цитокиндер эстафеталық принцип бойынша әрекет етеді; цитокиннің жасушаға әсер етуі оның басқа да цитокиндерді бөлуін тудырады (цитокиндік каскад). Цитокиндердің ерекшелігі: 1) биологиялық әсерінің жоғары сәйкестілігі, мысалы, ИЛ-2, ИЛ-4, ИЛ-7, ИЛ-12, ИЛ-15, ИЛ-18 үшін; 2)тұқымдастығының ИЛ-10 (ИЛ-19 – ИЛ-25) жаңа интерлейкиндер тән цитокиндердіңжекеленген топтарының бір ғанарецепторлық суббірлігімен әрекеттесу қабілеттілігі.
Цитокиндердің әсер ету механизмдерін интракинді, аутокринді, паракринді және эндокринді деп ажыратады.
Интракринді механизм – цитокиндердің жасуша-продуцент ішіндегі әсері;цитокиндердің өзіне тән жасушаішілік рецепторлармен байланысуы.
Аутокринді механизм – бөлінген цитокиннің өзін бөлген жасушаға әсері. Мысалы, моноцит/макрофагтар үшін белсендіруші аутокринды факторлар болып интерлейкин-1, -6, -18, ІНФ-α саналады (36-сурет).
Паракринді механизм – цитокиндердің жақын орналасқан жасушалар мен тіндерге әсері. Мысалы, макрофагтан бөлінген ИЛ-1, -6, -12 және -18, ІНФ-α антиген мен макрофагтың МНС-нтанитын Т-хелперді (Тһ0) белсендіреді.
Эндокринді механизм – цитокиндердің жасуша-продуценттен қашықтықтан әсер етуі. Мысалы, ИЛ-1, -6 және ІНФ-α ауто- және паракринді әсерден басқа, дистантты иммундыреттеуші, пирогенді, гепатоциттермен жедел фаза нәруыздарын өндіру индукциясына, уытты-септикалық жағдайлардағы мультиағзалық зақымдану және интоксикация белгілері болуы мүмкін.
Интерлейкиндер(ИЛ) – лейкоциттер арасындағы жасушааралық байланысқа жауапты цитокиндер. 25-тен көп интерлейкиндер анықталған. Жаңа ИЛ-19 – ИЛ-27интерлейкиндеріидентификафияланған.
ИЛ-1 макрофагтардан бөлінеді және аз көлемде дендритты жасушалардан, эндотелиоциттерден, фибробластардан, ТЖ, кератиноциттерден, кейбір Тһ2 клондарынан өндіріледі. Ол Т-хелпермен ИЛ-2 өндірілуін белсендіреді, Т-лимфоциттерде ИЛ-2 рецепторының пайда болуына әсерін тигізеді, В-лимфоциттің пісіп жетілуіне әсер етеді, МНС молекулаларыныңтүзілуін және гепатоциттермен жіті фазалынәруыздарының түзілуін белсендіреді. Нейтрофилдер мен ТЖфункциясын жоғарылатады. Қабынуалды және пирогенді әсер көрсетіп, иммундық, нерв және эндокриндік жүйелердің өзара байланысын қамтамасыз етеді.
ИЛ-2 Т-лимфоциттерден, негізінен Тһ1-мен,сонымен қатар 1 реттегіцитоуытты лимфоциттермен (СD8+) өндіріледі. Тһ1 және Т-жендеттердің дифференцировкасынбелсендіріп,ТЖ-ді жәнеВ-лимфоциттермен иммуноглобулиндердің синтезделуін ынталандырады.
ИЛ-3 Т-лимфоциттермен және бағаналы жасушалармен өндіріледі. Бағаналы және бастапқы гемопоэтикалық жасушалардыңөсу факторы болып табылады.
ИЛ-4 Тһ2, В-лимфоциттермен және мес жасушалармен өндіріледі. Тһ0-діңТһ2 бағытында дифференцировкасын және В-лимфоциттермен иммуноглобулиндердің синтезделуін ынталандырады. Цитоуытты лимфоциттердің,ТЖ-дің генерациясын,сонымен қатар ИФН-γ өндірілуін және макрофагтардың ісікке қарсы белсенділігін тежейді.
ИЛ-5 Тһ2-мен синтезделеді. Белсенген В-лимфоциттердің пролиферациясымен саралануына ықпал жасайды, IgА өндірілуін күшейтеді,эозинофилдерді белсендіреді.
ИЛ-6 макрофагтардан, Т-және В-лимфоциттерден өндіріледі. Тимоциттердің, В-лимфоциттердің пролиферациясын ынталандырып, цитоуытты лимфоциттердің, гранулоцит пен макрофагтардың ізашарларын белсендіреді. Гепатоциттерден жедел фаза нәруыздарының пайда болуын белсендіріп,қабынуалды әсер көрсетеді, иммундық, нерв және эндокриндік жүйелердің өзара байланысын қамтамасыз етеді.
ИЛ-7 сүйек кемігімен тимустың арқаулық жасушаларынан өндіріледі. Ізашар В- және ізашар Т-лимфоциттерінің өсу факторы болып табылады.
ИЛ-8 моноциттермен, макрофагтармен, фибробласттармен өндіріледі. Қабыну ошағына нейтрофилдермен базофилдердің миграциясын және олардың дегрануляциясын,супероксидті радикал бөлінуін тудырады. Ангиогенезді ынталандырады.
ИЛ-9 негізінен Т-лимфоциттермен өндіріледі. Т-лимфоциттердің пролиферациясын ынталандырады, мес жасушаларды белсендіреді, эритропоэтиннің әсерін күшейтеді.
ИЛ-10 Тһ2-мен, сонымен қатар екінші реттегі цитоуытты Т-лимфоциттермен және макрофагтармен өндіріледі. В-лимфоциттердің пролиферациясын және саралануынынталандырып, Тһ1-жасушаларымен ИЛ-2 және ИФН-γ түзілуін тежейді, жасушалық иммундықжауапты және қабынуалды цитокиндердіңөндірілуін тежейді.
ИЛ-11 сүйек кемігінің арқаулық жасушаларынан өндіріледі. Гемопоэз ізашарларының саралануы мен бөлінуін, мегакариоциттердің клон түзуін ынталандырып, шеткері қанда тромбоциттер мен эритроциттер санын жоғарылатады.Қабынуалды цитокиндерініңөндірілуін тежейді.
ИЛ-12 моноциттер мен макрофагтарданжәне аз көлемде В-лимфоциттер мен дендритті жасушалардан өндіріледі. Тһ (Тһ0 ꞊›Тһ1), Т-жендет, ТЖсаралануын және өсуін ынталандырады. Т-лимфоциттермен және ТЖ-ден ИФН-γ өндірілуін индукциялайды. Тһ1-дің апоптозын, IgЕ түзілуін тежейді. ИЛ-4-пен бірігіп Тһ1 және Тһ2 тепе-теңдігін реттейді.
ИЛ-13 Тһ2 және мес жасушалармен түзіледі.В-лимфоциттердіңсаралануы мен өсуін белсендіреді, моноцит/макрофаг қызметін, әсіресеқабынуалды цитокиндер өндірілуін тежейді.
ИЛ-14 негізінен Т-лимфоциттермен өндіріледі.В-лимфоциттердің пролиферациясын күшейтеді және иммуноглобулиндердің өндірілуін тежейді.
ИЛ-15 моноцитттермен, эпителиоциттермен және тегіс бұлшықеттік жасушалармен өндіріледі.ИЛ-15 Т-лимфоциттерге әсері бойынша ИЛ-2-ге ұқсас, ол ИЛ-2 рецепторымен өзіне тән түрде байланысу қабілетімен түсіндіріледі. В-лимфоциттер мен ТЖ-ді белсендіреді.
ИЛ-16 эозинофилдерден және СD8+ Т-лимфоциттермен өндіріледі. СD4+ Т- лимфоциттердің хемотаксисін белсендіреді.
ИЛ-17 белсендірілгенСD4+ Т- лимфоциттермен, сонымен қатар ТЖ жасушаларымен өндіріледі. Цитокиннің негізгі нысана-жасушалары болып эпителиоциттер, эндотелиоциттер және фибробласттар саналады. Ол ИЛ-6, ИЛ-8, гранулоцитарлы КСФ, простогландин Е2 өндірілуін, ІСАМ-1 экспрециясын күшейтеді, фибробласттардың белсенділігін ынталандырады.
ИЛ-18 белсенген макрофагтармен, сонымен қатар гепатоциттермен өндіріледі. Т-лимфоциттермен ИФН-γ, макрофагтармен ИЛ-1, ИЛ-8 және ІНФ өндірілуін белсендіреді. Сондай-ақ, ол ТЖ-ті белсендіреді.
Ісік некрозының факторлары (ІНФ).Ісік некроздаушы факторларнемесе ІНФ-α; лимфотоксиндер, немесе ІНФ-β деп ажыратылады. ІНФ -α макрофагтармен, сонымен қатар мес жасушалармен және лимфоциттермен өндіріледі. Ол уытты шок пен кахексияның (ескі атауы −кахектин)дамуына әкеледі, жедел фазалық нәруыздарды индукциялап, ангиогенезді ынталандырады. Апоптозды индукциялауы мүмкін. Бірқатар ісіктердің гемморагиялық некрозын тудыруы мүмкін. ІНФ-β Т- және В-лимфоциттерден өндіріледі және әсері ІНФ-α ұқсас.
Интерферондар (ИФН) – вирустықжұқпаларғажәне басқа да ынталандыруларға жауап ретінде жасушалардан өндірілетін гликопротеиндер. Вирустың басқа жасушалардағы репродукциясын тежейді және иммундық жүйе жасушаларының өзара әрекетіне қатысады. Интерферондардың екі серологиялық тобын ажыратады: І типі – ИФН-α және ИФН-β, ІІ типі− ИФН-γ. І типті интерферондар вирусқа қарсы және ісікке қарсы әсерін көрсетсе, ал ІІ типті интерферондар спецификалық иммундық жауапты және бейспецификалық резистенттілікті реттеп отырады.
ИФН-α және ИФН-β құрылысы мен жасуша-продуцентібойынша ерекшеленсе де, әсер ету механизмдері бірдей. Қалыпты жағдайда ИФНα – мононуклеарлы фагоциттермен (осыдан бір атауы – “лейкоцитарлықИФН”), ал ИФН-β фибробласттармен (“фибробласттық ИФН”)өндіріледі. Микробтардың әсерінен олар көптеген жасушалардан бөлінеді. ИЛ-1-діңпирогенді әсеріненжәне дене қызуының жоғарлауына байланысты жасушааралық сұйықтықтардың рН-ың төмендеуі ИФН-дың өндірілуін күшейтеді. І типті ИФН−дың қорғаныштық әсері құрамында интерферондары бар жасушалардан өндірілетін олигоаденилатсинтетазаның әсерінен РНК немесе ДНК репликациясының тежелуі нәтижесінде іске асырылады.
І типті ИФН-дар сау жасушалармен байланысып, оларды вирустардан қорғайды. І типті ИФН-ң вирустарға қарсы әсер етуі аминқышқылдардың, мысалы,триптофанныңөндірілуіне кедергі жасау арқылы оның жасушалық пролиферациясын тежеу мүмкіндігімен байланысты. Бұл механизм, сондай−ақ кейбір ісіктердің бағдарламаланған жасуша өлімін индукциялау қабілеті І типті ИФН-дың ісікке қарсы әсерінің негізінде жатыр. Сонымен қатар, І типті ИФН-дар ТЖ-дің нысана-жасушаларға, соның ішінде трасформацияланған жасушаларғализизтік әсерін күшейтеді, МНС І анитигендерінің экспресиясын индукциялайды және керісінше сол бір МНС ІІ антигендерінің қалыптасуын тежейді.
ИФН-γ (иммундық ИФН) Т-лимфоциттер мен ТЖ-мен өндіріледі. Т- және В-лимфоциттердің, моноцит/макрофагтар және нейтрофилдердің белсенділігін ынталандырады. МНС І, МНС ІІ молекуласының экспрециясын күшейтеді. Тһ0-дің Тһ1-ге саралануын ынталандырады. Иммундық ИФН өзінің антагонисті ИЛ-4-пен бірге Тһ1/Тһ2 тепе-теңдігін ұстап тұрады. Оған қоса, бірқатар барлық қалыпты, сондай-ақ кейбір трансформацияланған және инфекцияланған жасушалардың апоптозын реттейді. Осылайша, ол белсендірілген макрофагтардың, кератиноциттердің, гепатоциттердің, сүйек кемігі жасушаларының, эндотелиоциттердің апоптозын индукциялайды және шеткері моноциттер мен герпес−жұқтырған нейрондардың апоптозын тежейді.
Колониястимулдаушы факторлар (КСФ) – сүйек кемігіндегі бағаналы жасушалар мен қан жасушаларының ізашарларының бөлінуін, саралануын реттеуші цитокиндер. Сонымен қатар, олар сүйек кемігінен тыс кейбір жасушалардың саралануы мен функциялық белсенділігін ынталандыра алады.
Гранулоцитарлық КСФ (Г-КСФ) көбіне макрофагтармен, сонымен қатар фибробласттармен өндіріледі. Бағаналы жасушалардың бөлінуін және саралануын белсендендіреді, нейтрофилдер мен эозинофилдердің белсенділігін күшейтеді.
Макрофагалды КСФ (М-КСФ) моноциттермен, аз мөлшерде эндотелиалды жасушалармен және фибробласттармен өндіріледі. Сүйек кемігіндегі макрофагтардың ізашарларының пролиферациясын белсендендіреді.
Гранулоцитарлы-макрофагалды КСФ (ГМ-КСФ) макрофагтармен және Т-лимфоциттермен, сонымен қатар фибробласттармен және эндотелиоциттермен өндіріледі. Гранулоциттердің ізашарлары мен макрофагтардың бөлінуін және саралануын белсендіреді, макрофагтар мен гранулоциттерді, Т-жасушалардың пролиферациясын белсендендіреді. Қан өндіретін ізашарлардың антигентаныстырушы дендритті жасушаларғасаралануын ынталандыруға қатысады.
Трансформациялайтын
өсу факторы- (ТӨФ-
(ТӨФ- )
–полифункционалды
өсу факторы, оған сондай-ақ фибробласттардың,
тромбоциттердің, эндотелийдіңөсу
факторлары, инсулин тәрізді өсу факторы,
эпидермалды өсу факторы және т.б. жатады.
ТӨФ-
)
–полифункционалды
өсу факторы, оған сондай-ақ фибробласттардың,
тромбоциттердің, эндотелийдіңөсу
факторлары, инсулин тәрізді өсу факторы,
эпидермалды өсу факторы және т.б. жатады.
ТӨФ- көптеген жасушалардан (көбінесе өндіруші
− макрофагтар), соның ішінде кейбір
ісік жасушаларымен өндіріледі. ТӨФ-
көптеген жасушалардан (көбінесе өндіруші
− макрофагтар), соның ішінде кейбір
ісік жасушаларымен өндіріледі. ТӨФ- моноцит/макрофагтар үшін олардың
цитоуытты және цитокин−өндіруші
белсенділігін, сонымен қатар беткейіндегі
МНС молекулаларының экспрессиясын
төмендетуші − қуатты деактивтендіруші
фактор. Бұл қатынаста ол басқа
макрофаг−деактивтендіруші цитокиндермен
(ИЛ-4, -10 және -13) бірігіп әсер етеді.
Моноцитарлы/макрофагалды қатардың
жасушаларынан нитроқосылыстардың,
реакциялық-қабілетті радикалдардың
және қабынуалды цитокиндерінің өндірілуін
төмендететін қасиетіне байланысты
ТӨФ-
моноцит/макрофагтар үшін олардың
цитоуытты және цитокин−өндіруші
белсенділігін, сонымен қатар беткейіндегі
МНС молекулаларының экспрессиясын
төмендетуші − қуатты деактивтендіруші
фактор. Бұл қатынаста ол басқа
макрофаг−деактивтендіруші цитокиндермен
(ИЛ-4, -10 және -13) бірігіп әсер етеді.
Моноцитарлы/макрофагалды қатардың
жасушаларынан нитроқосылыстардың,
реакциялық-қабілетті радикалдардың
және қабынуалды цитокиндерінің өндірілуін
төмендететін қасиетіне байланысты
ТӨФ- -ны
қабынуға қарсы цитокиндерге жатқызады.
Бірақ кейбір жағдайларда ол қабынуалды
әсер беруі мүмкін.
-ны
қабынуға қарсы цитокиндерге жатқызады.
Бірақ кейбір жағдайларда ол қабынуалды
әсер беруі мүмкін.
Қалыпты
және трансформацияланған жасушалардың
бағдарламаланған жасушалық өлім үдерісін
реттейді. ТӨФ- Тһ1 апоптозын тежейді, ал ИЛ-2-мен бірге
Тһ2 апоптозын тежейді. Иммундық жүйе
жасушаларының апоптозын тежей отырып,
жад жасушасының генерациясында маңызды
рөл атқаруы мүмкін. Осы және басқа да
кейбір өсу факторларының артық
белсенділігі гиперпролиферативті
үдеріске, яғни гломерулонефрит, терінің
склероздануы, бауыр циррозы және т.б.,
сонымен қатар ісіктің прогрессивті
өсуіне әкелуі мүмкін. ТӨФ-
Тһ1 апоптозын тежейді, ал ИЛ-2-мен бірге
Тһ2 апоптозын тежейді. Иммундық жүйе
жасушаларының апоптозын тежей отырып,
жад жасушасының генерациясында маңызды
рөл атқаруы мүмкін. Осы және басқа да
кейбір өсу факторларының артық
белсенділігі гиперпролиферативті
үдеріске, яғни гломерулонефрит, терінің
склероздануы, бауыр циррозы және т.б.,
сонымен қатар ісіктің прогрессивті
өсуіне әкелуі мүмкін. ТӨФ- −
неопластикалық аурулар кезінде
иммуносупрессор болып табылатын
медиаторлардың бірі.
−
неопластикалық аурулар кезінде
иммуносупрессор болып табылатын
медиаторлардың бірі.
Хемокиндер – жасушалар хемотаксисіне жауапты төменмолекулалық цитокиндер (шамамен 40-MIP, MCP, RANTES, ИЛ-8 және т.б.), (қабыну ошағына лимфоциттер мен лейкоциттерді тартады). Лейкоциттердің қозғалғыштығын реттейді.