

4.2.3. Влияние pH
Многие клеточные культуры растут в достаточно узком диапазоне pH.
Влияние pH:
на субстрат (напр, аминокислоты);
на ферменты клетки.
Кинетика переноса ионов водорода через мембрану клетки:
![]() (4.17)
(4.17)
В условиях равновесия:
![]() (4.18)
(4.18)
4.3. Замедление скорости роста
4.3.1. Лимитирование по субстрату
![]() (линейная
связь) (4.19)
(линейная
связь) (4.19)
![]() (4.20)
(4.20)
Тогда
 (4.21)
(4.21)
Решение:
![]() (4.22)
(4.22)
Уравнение (4.22) называют интегральным уравнением Моно. Предел роста:
![]() (4.23)
(4.23)
4.3.2. Ингибирование избытком субстрата
 (4.24)
(4.24)
![]() (4.25)
(4.25)
 (4.26)
(4.26)
 (4.27)
(4.27)
4.3.3. Ингибирование субстратом в условиях истощения по субстрату
 (4.27)
(4.27)
Решение:
![]()
4.3.4. Ингибирование продуктами
 (4.28)
(4.28)
![]() (4.29)
(4.29)
![]() (4.30)
(4.30)
![]() (4.31)
(4.31)
 (4.32)
(4.32)
Уравнение (4.32) преобразуется к виду
 (4.33)
(4.33)
Решение:
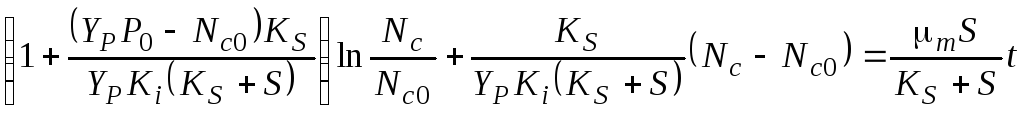 (4.34)
(4.34)
4.3.5. Период индукции
1) Трансформация предсубстрата в субстрат.
![]() (4.35)
(4.35)
![]() (4.36)
(4.36)
![]() (4.37)
(4.37)
Решение:
![]() (4.38)
(4.38)
где
![]() (4.39)
(4.39)
![]() (4.40)
(4.40)
Период индукции:
![]() (4.41)
(4.41)
2) Адаптация.
![]() (4.42)
(4.42)
![]() (4.43)
(4.43)
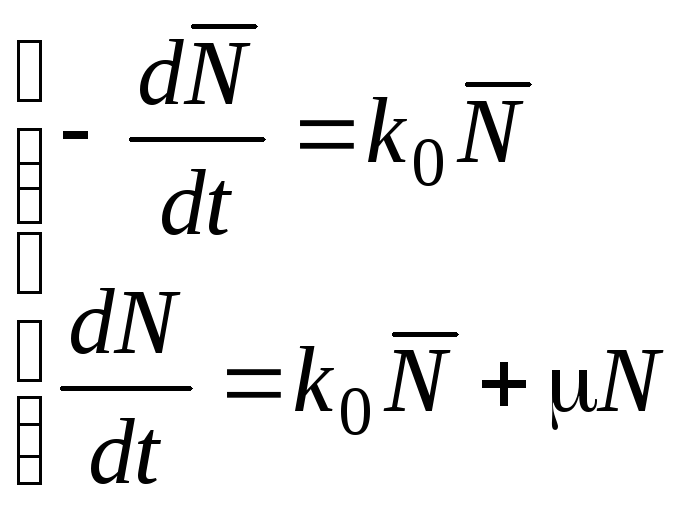 (4.44)
(4.44)
![]() (4.45)
(4.45)
![]() (4.46)
(4.46)
Период индукции:
![]() (4.47)
(4.47)
Расход ингибитора роста.
![]() (4.48)
(4.48)
![]() (4.49)
(4.49)
![]() (4.50)
(4.50)
Дифференциальное уравнение для скорости роста культуры:
![]() (4.51)
(4.51)
Период индукции:
![]() (4.52)
(4.52)
4.4. Старение клеток и апоптоз
Апоптоз – совокупность процессов “программируемого” старения и гибели клеток. Некроз – гибель клеток за счет случайных событий или под действием внешних токсинов.
1961 г., Л. Хейфлик: предел клеточного деления для фибробластов человека (50 делений).
Бактериальные клетки и клетки одноклеточных организмов могут делиться бесконечно; клетки многоклеточных организмов имеют предел деления (существует максимальное количество делений).
Концепция “программируемого” старения: ДНК полимераза не способна реплицировать “хвосты” 3’ конца матрицы ДНК (несколько нуклеотидов на 3’ конце). Для предотвращения укорачивания ДНК фермент теломераза синтезирует на концах ядерной ДНК многократно повторяемый гексануклеотид TTAGGG (теломера). Таким образом, для преодоления укорачивания генома и старения клетка должна активировать теромеразный ген и экспрессировать большое количество теломеразы.
4.4.1. Выражение для удельной скорости клеточного роста в экспоненциальной фазе
 (4.53)
(4.53)
Предположения:
- скорость синтеза ДНК пропорциональна концентрации S’ и характеризуется константой скорости R;
- скорость биосинтеза лимитирующего фермента и белков репликационного комплекса намного выше скорости синтеза ДНК, так что можно считать E=const.
Тогда:
 (4.54)
(4.54)
В квазистационаре:
![]() (4.55)
(4.55)
следовательно
![]() (4.56)
(4.56)
Квазистационарная концентрация синтезируемой ДНК:
![]() (4.57)
(4.57)
Репликационный процесс заканчивается, когда количество синтезируемой ДНК равно базовому количеству ДНК:
![]() (4.58)
(4.58)
Отсюда находим время репликации:
![]() (4.59)
(4.59)
И удельная скорость роста клеточной культуры в экспоненциальной фазе:
![]() (4.60)
(4.60)