

4.2.20. Проблема управляемого культивирования
В последние десятилетия XX в. одной из интенсивно развивающихся проблем микробиологии стала проблема управляемого культивирования. Теоретической основой возникновения и развития этого направления явилось детальное изучение большого разнообразия физиологических потребностей микроорганизмов. Результаты физиологических исследований позволили с помощью соответствующих условий культивирования регулировать ход бродильных и ферментационных процессов, накапливать микробную массу с заданными свойствами, воздействовать на ход микробиологических процессов.
Разработка проблемы управляемого культивирования восходит к исследованиям более раннего периода (20-е годы), направленным на поиск методов изменения хода обмена веществ микроорганизмов с помощью разнообразных факторов внешней среды. Впервые на принципиальную возможность управлять развитием культуры с помощью условий среды указал Г. Клебс (1905), которому удалось таким образом регулировать развитие грибов и водорослей. В 50-е годы XX в. учеными разных стран была разработана теория роста и развития микробов (М. Стефенсон, И. Гунсалус, Н. Д. Иерусалимский, Ж. Моно, В. Шеффер, Р. Финн и др.). Были разработаны теоретические основы управления ростом микробов, опирающиеся на принцип минимума («узкого места»), т.е. использование функции лимитирующего вещества (Ж. Моно, А. Новик, Л. Снилард и др.), а также на регулирование величины популяции и накопления продуктов жизнедеятельности. Полученные данные позволили найти практические методы управляемого культивирования микробов, в том числе метод непрерывного, или проточного, культивирования, теоретическое обоснование которого было дано Ж. Моно (рис. 4.2.44) в 1950г.

Рис. 4.2.44. Жак Люсьен Моно (Jacques Lucien Monad, 1910–1976)
Метод состоит в подаче в культиватор с постоянной скоростью питательной среды и непрерывном выводе с такой же скоростью культуральной среды с бактериями. Точный контроль за непрерывным культивированием микробов осуществляется в аппаратах типа турбидостата, обеспечивающего контроль за плотностью биомассы микроба, либо типа хемостата, позволяющего контролировать скорость увеличения концентрации лимитирующего фактора (рис. 4.2.45).
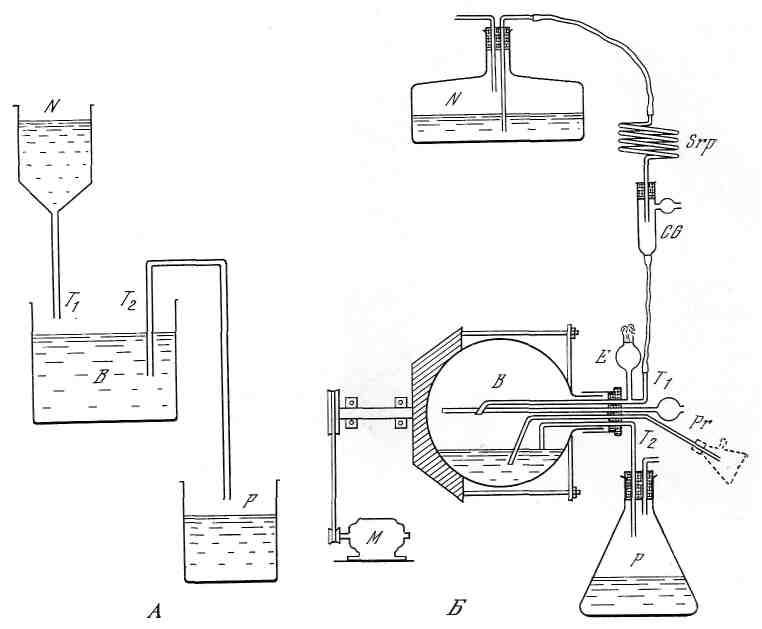
Рис. 4.2.45. Турбидостат Моно (Б) и схема проточной установки (А) по Моно (1950)
Значительное повышение точности физиологических исследований за счет обеспечения постоянства условий среды и состояния культуры определило широкое использование метода непрерывного культивирования в микробиологии.
4.2.21. Основные этапы развития генетики микроорганизмов
В 40—50-е годы XX в. из общей микробиологии выделилось самостоятельное направление — генетика микроорганизмов — с наиболее важным разделом — генетикой бактерий. Основой исключительно быстрого развития этой области исследований послужили доказательства мутационной природы изменчивости микроорганизмов, обнаружение у них различных форм генетической рекомбинации и роли ДНК в их наследственных свойствах, достоинства микроорганизмов как объектов генетических исследований. Генетика микроорганизмов приобрела особое значение в связи с решением кардинальных проблем молекулярной биологии. Исследования структуры ДНК, ее роли в процессах биосинтеза белка и регуляции внутриклеточных обменных процессов, разработка проблем направленной изменчивости и специфичности мутагенеза ведутся в основном на микроорганизмах.
До оформления генетики микроорганизмов в самостоятельную отрасль знания генетики и микробиологи работали разобщенно. Микробиологи были далеки от генетического истолкования наблюдавшихся ими фактов изменчивости и наследственности микробов, а генетики либо вообще не интересовались бактериологией, либо скептически относились к возможности применения генетических принципов к таким примитивно организованным формам жизни, какими им представлялись бактерии.
Первый этап в развитии генетики бактерий составили исследования, экспериментально доказывающие сходство природы и механизмов передачи потомству наследственных признаков у бактерий и высших организмов.
В период с 1910 по 1940 г. были описаны различные виды наследственной изменчивости. В 1921 г. французский микробиолог П. де Крюи описал изменение ряда свойств у одной из патогенных бактерий, возникающее, по его мнению, в результате расщепления признаков под влиянием неблагоприятных условий среды. Этот вид изменчивости он назвал диссоциацией. В 1925 г. в Советском Союзе Г.А. Надсон и Г.С. Филиппов впервые получили мутационные (по их терминологии, сальтационные) стойкие изменения признаков у дрожжевых и плесневых грибков, подвергнув их облучению рентгеновыми лучами. Это был первый случай индуцированной мутации, вызванной физическим мутагенным фактором. Первые данные о характере мутаций позволили провести параллель между природой изменчивости у бактерий и высших организмов. Это стало возможным также после того, как были разработаны методы, которые дали возможность отличать изменения отдельной клетки от изменчивости целой популяции.
Причины различий в объяснении фактов изменчивости у микробов носили преимущественно методологический характер. Из-за отсутствия единой методики эксперимента данные разных авторов оставались несопоставимыми. Трудности в разграничении фенотипа и генотипа приводили зачастую к отрицанию различий между адаптацией и мутацией. Случаи морфологической изменчивости, требующие длительного наблюдения, нередко объясняли наличием сложных циклов развития. Длительное время отсутствовали и единые методы генетического анализа, в частности, принцип отбора мутантов. Значительное упорядочение представлений о природе изменчивости у микробов было связано с разработкой методов генетического анализа у высших организмов и утверждением представлений о сходстве механизмов изменчивости и наследственности у всех живых организмов.