

4.2.23. Изучение трансформации, трансдукции, конъюгации и лизогенной конверсии
В настоящее время трансформацией называют процесс переноса информации с помощью ДНК от клетки-донора к клетке-реципиенту и замещения в последней в результате рекомбинации специфической последовательности нуклеотидов генома. Термин «трансформация» появился в 1928—1934 гг., когда было обнаружено, что некоторые штаммы бактерий, выращенные в присутствии убитых клеток или в культуральных фильтратах и экстрактах из других родственных штаммов, могли приобретать некоторые свойства этих штаммов (рис. 4.2.46). Сущность этого явления была выяснена в 1944 г. О. Эвери, К. Мак-Леодом и М. Мак-Карти, установившими, что трансформирующим агентом является высокомолекулярная ДНК.
Дальнейшее изучение трансформации проводилось при учете в качестве маркеров различных признаков: капсулообразования (для пневмококков), устойчивости к антибиотикам, потребности в факторах роста (X. Раппапорт, 1959; Л. Толмач, Р. Херриотт, 1962; и др.). Исследования с использованием последних двух маркеров дали возможность детально разработать проблемы множественной трансформации и «сцепления» различных генов. Важное место среди этих исследований заняли работы японских ученых X. Иошикавы и Н. Сеока, которые в 1963 г. разработали метод картирования сцепленных генов у Bacillus subtilis.
Способность ДНК проникать в клетку-реципиент зависит как от природы самой ДНК, так и от физиологического состояния клетки-реципиента. Трансформирующей ДНК могут быть только высокомолекулярные двухцепочечные фрагменты, при этом проникать в бактериальную клетку может ДНК, выделенная из разных биологических источников, но включаться в геном — только ДНК с определенной степенью гомологичности. После того как экзогенный фрагмент ДНК, проникший в клетку, нашел гомологичный фрагмент ДНК клетки-реципиента, между ними происходит генетический обмен.

Рис. 4.2.46. Схема эксперимента Гриффита (по Стенту): а — мышь, которой введена культура патогенного капсулированного штамма S пневмококов, погибает; б — мышь, которой введена культура непатогенного бескапсульного R—мутанта нормального S—штамма, не погибает; в — мышь, которой введена культура S—штамма, убитого предварительно нагреванием, не погибает; г — мышь, которой введена смесь живой культуры R—мутанта и убитой нагреванием культуры нормального S—штамма, погибает; в этом случае присутствие убитых нагреванием S—бактерий вызвало трансформацию живых R—бактерий, в результате чего у них восстановилась способность к образованию капсулы и патогенность.
Открытие лизогении связано с именами Ж. Борде и М. Чуке (1921), которые положили начало изучению роли бактериофага в осуществлении бактериальных рекомбинаций (рис. 4.2.47). Данное ими определение лизогении как наследственной способности бактерий спонтанно продуцировать бактериофаг при отсутствии экзогенного заражения сохранилось и по сегодняшний день.

Рис. 4.2.47. Встраивание ДНК фага X в хромосому Е. coliв неэкспрессируемом состоянии, которое может поддерживаться посредством репликации в течение многих поколений. В результате некоего события, играющего роль пускового механизма, вирусный геном может начать экспрессироваться с образованием фаговых частиц и последующим лизисом клеток.
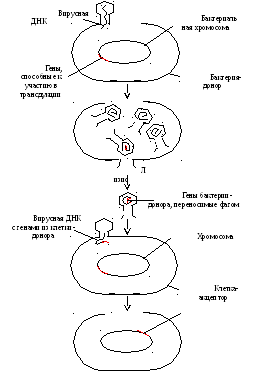
Рис. 4.2.48. Трансдукция. Если бактериальная клетка заражена некоторыми ДНК-содержащими фагами, то небольшая часть ее хромосомы может ковалентно присоединиться к фаговой ДНК, реплицироваться вместе с ней и таким образом встраиваться в ДНК дочерних фаговых частиц. Когда такие частицы заражают другую клетку, фаговая ДНК приносит в эту клетку участок хромосомы первой клетки.
На основе исследований Ф. Бернета, М. Дельбрюка и особенно А. Львова, проливших свет на истинную природу явления лизогении, стало возможным открытие Дж. Ледербергом и Н. Циндером (1952) нового способа переноса генетической информации с помощью бактериофага, названного ими трансдукцией (рис. 4.2.48). Трансдукция оказывается возможной, если в процессе размножения фага одна из частиц случайно захватит фрагмент бактериальной хромосомы, как правило, содержащий очень небольшое число генов. Когда такая фаговая частица заражает бактерию-реципиент, бактериальная ДНК проникает в клетку таким же путем, как фаговая. Между трансдуцированной бактериальной ДНК и гомологичным участком бактериальной хромосомы может произойти обмен, и как следствие его возникают рекомбинанты, несущие небольшую часть генетического материала клетки-донора. Передача признаков с помощью фагов показана для бактерий, принадлежащих к разным родам. Использование разных форм трансдукции способствовало решению многих сложных генетических проблем.
В 50-х годах Б. Хейс установил, что при бактериальном скрещивании наблюдается полярность, причем один из партнеров каждой конъюгирующей пары является донором, или «самцом», а другой — реципиентом, или «самкой», т.е. партнеры, участвующие в конъюгации, гетероталличны. Им были выделены два половых типа — F+ (донорные, или мужские штаммы) и F- (реципиентные, или женские штаммы). В 1956 г. Дж. Ледерберг представил прямые микроскопические доказательства образования конъюгационных пар в смешанных культурах рекомбинирующих штаммов.
При конъюгации, для которой необходим непосредственный контакт между бактериальными клетками, осуществляется направленный перенос генетического материала от клетки-донора в клетку-реципиент (рис. 4.2.49). Как правило, в клетку-реципиент переносится только часть генетического материала клетки-донора, в результате чего образуется неполная зигота, или мерозигота, содержащая часть генома донора и полный геном клетки-реципиента. Участки перенесенной от донора ДНК находят гомологичные участки в молекуле ДНК реципиента, между которыми происходит генетический обмен. В результате часть донорной ДНК встраивается (интегрируется) в геном реципиента, а соответствующая часть реципиентной ДНК из него исключается.
Наконец, еще один путь переноса генетического материала у прокариот осуществляется с помощью плазмид определенного типа, обладающих генами, обеспечивающими эту возможность. Такие плазмиды помимо переноса собственного генетического материала могут обеспечивать перенос хромосомных генов, плазмид, не обладающих способностью к самостоятельному переносу, а также осуществлять передачу транспозонов из плазмиды в хромосому или другую плазмиду.
Все известные способы передачи генетической информации с помощью плазмид создают огромные возможности для интенсивных генетических обменов между клетками различных бактерий. Плазмидам и другим нехромосомным генетическим элементам принадлежит основная роль в передаче генетической информации «по горизонтали». Можно предположить, что в природе любая генетическая информация может быть перенесена в любую клетку прокариот, если не прямо, то через посредников. Подтверждением этого могут служить данные по введению с помощью сконструированной плазмиды в бактериальную клетку эукариотной ДНК и ее репродукции там.

Рис. 4.2.49. Конъюгация бактерий. Обычно бактерии размножаются вегетативным путем, с помощью простого роста и деления. Однако у некоторых бактерий иногда происходит половая конъюгация, в ходе которой часть одной из цепей (или вся цепь) хромосомы донорной клетки переносится через пиль – длинный соединительный канал – в реципиентную клетку того же вида.
Как редкое событие, происходящее с частотой 10–4—10–7, плазмиды или отдельные гены, входящие в их состав, могут включаться в бактериальную хромосому. Поскольку ДНК плазмиды и бактериальной клетки не имеют одинаковых нуклеотидных последовательностей, т.е. не являются гомологичными, рекомбинация между ними происходит не по механизму обмена, а по механизму встраивания. Рекомбинации такого типа происходят также с участием транспозонов и IS-элементов при их перемещении (транспозиции) в пределах хромосомы. Встраивание плазмид и мигрирующих элементов помимо того, что приводит к введению в хромосому дополнительного генетического материала, может вызывать перестройку бактериального генома: нарушать целостность генов или регуляцию их функционирования, т.е. вызывать мутации.