

8) Окисление высокомолекулярных кислот жирного ряда
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концов проходит через стадию масляной кислоты. После очередного β-окисления масляная кислота становится ацетоуксусной. Последняя затем гидролизуется до двух молекул уксусной кислоты.
Наиболее распространенным механизмом ассимиляции жирных кислот микроорганизмами является β-окисление. Оно заключается в последовательном отщеплении двух углеродных атомов.
Следует рассмотреть следующие основные этапы β-окисления:
1)Активация жирной кислоты;
2)Транспорт ацильной группы;
3)Последовательность реакций β-окисления;
4)Энергетика окисления жирных кислот.
Реакция окисления жирной кислоты происходят только после превращения ее в активированную высокоэнергетическую форму - ацил-КоА. Этот процесс требует затраты одной молекулы АТФ, присутствия коэнзима А и ионов Mg2+; катализирует превращение свободной жирной кислоты в активированную форму – фермент ацил – КоА-синтетаза (тиокиназа):

Затем происходит дегидрирование, катализируемое FAD-зависимой ацил-КоА-дегидрогеназой,приводит к образованию α,β-ненасыщенного ацил-SКоА. Фермент обладает стереоспецифичностью, поэтому в результате этой реакции образуется только транс-изомер (транс-еноил- КоА):

Образовавшийся транс-еноил-КоА подвергается гидратации при действии фермента еноил-КоА-гидратазы.В результате образуется L- β-гидроксиацил- КоА:
Затем вновь происходит дегидрирование, в процессе которой образовавшийся β-гидроксиацил-КоА дегидрируется в β-положение.Эту реакцию катализирует NAD-зависимая β-гидроксиацил-КоА-дегидрогеназа:
На заключительном этапе расщепления β-кетоацил-КоА с помощью еще одной молекулы коэнзима А образуется укороченная на два углеродных атома ацил-КоА и двухуглеродный фрагмент в виде ацетил-КоА. Реакция катализируется ацетил-КоА-ацилтрансферазой (тиолазой):
Образующийся укороченный ацил-КоА вновь вступает в следующий цикл β-окисления, начиная с первой реакции дегидрирования, и происходит повторное превращение этого ацил-КоА в цикле, состоящем из четырех реакций. Такой процесс β-окисления протекает до образования четырехуглеродного соединения – ацетоацетил-КоА. Последняя реакция этого соединения приводит к образованию двух молекул ацетил-КоА и тем самым завершает распад жирной кислоты по механизму β-окисления.
Молекулы ацетил-КоА, образовавшиеся из жирной кислоты, подвергается полному окислению до Н2О и СО2 в цикле трикарбоновых кислот.
Механизм окисления жирных ненасыщенных кислот аналогичен.
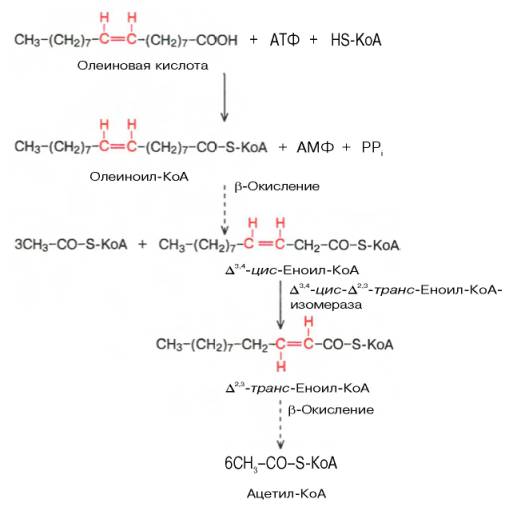
Двойные связи природных ненасыщенных кислот имеют цис-конфигурацию, в отличии от промежуточных продуктов β-окисления, имеющих транс-конфигурацию. К тому же последовательное укорачивание цепи часто приводит к образованию двойной связи в положении 3-4, гидротацию которой еноил-гидратаза не может осуществлять. В этом случае осуществляется реакция перемещения двойной связи от 3-го ко 2-му атому углерода, сопряженная с изменением конфигурации двойной связи. Реакцию катализирует 3,4-цис-2,3-транс-еноил-КоА-изомераза. Полиненасыщенные жирные кислоты одновременно подвергаются гидрированию под действием данного фермента.
Высокомолекулярные жирные кислоты (С14-С18) у некоторых микроорганизмов могут подвергаться α-окислению, т.е. последовательное отщепление одноуглеродных фрагментов от карбоксильного конца молекулы. Оно заключается в последовательном декарбоксилировании кислоты с образованием альдегида. Альдегид под действием НАД-зависимой дегидрогеназы окисляется с образованием соответствующей кислоты.
Рассмотрим энергетическую эффективность процесса β-окисления жирной кислоты, на примере пальмитиновой кислоты (C16H32O2).
При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5 х 7 = 35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле трикарбоновых кислот, дает 12 молекул АТФ, а 8 молекул ацетил-КоА дадут 12 х 8 = 96 молекул АТФ.
Таким образом, всего при полном β-окислении пальмитиновой кислоты образуется 35 + 96 = 131 молекула АТФ. С учетом одной молекулы АТФ, потраченной в самом начале на образование активной формы пальмитиновой кислоты (пальмитоил-КоА), общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты составит 131 – 1 = 130 молекул АТФ.
Таким образом жиры представляют большую энергетическую ценность, чем углеводы.