

4.1. Перелік основних термінів, параметрів, характеристик, які повинен засвоїти студент при підготовці до заняття.
|
Термін |
Визначення |
|
Мембрани |
Високопровідні структури клітини |
|
Фільтрація |
Масовий потік рідини через мембрану, що здійснюється внаслідок різниці гідростатичних тисків |
|
Дифузія |
Рух молекул розчиненої речовини з області з більшою в область з меншою її концентрацією |
|
осмос |
Рух розчинника із області з меншою концентрацію в область з більшою концентрацією |
4.2. Теоретичні питання до заняття.
Вкажіть основні структурні компоненти мембран.
Чим забезпечується механічна міцність мембран?
Які компоненти входять до складу мембранних фосфоліпідів?
Назвіть динамічні характеристики мембрани як рідкого кристалу.
Чому дорівнює товщина клітинної мембрани?
Що таке «кінки»?
Зміст теми
О сновні
групи речовин, з яких складаються
мембрани –це
фосфоліпіди й білки.
Крім них, у різних кількостях можуть
бути присутні вуглеводи, неорганічні
іони й вода, ліпіди інших груп, а згідно
деяких даних – і нуклеїнові кислоти.
сновні
групи речовин, з яких складаються
мембрани –це
фосфоліпіди й білки.
Крім них, у різних кількостях можуть
бути присутні вуглеводи, неорганічні
іони й вода, ліпіди інших груп, а згідно
деяких даних – і нуклеїнові кислоти.
Фосфоліпіди мембран в основному є фосфогліцеридами. Це складні ефіри трьохатомного спирту гліцерину, що містять у своєму складі 2 довгих вуглеводних радикала жирних кислот (неполярна гідрофобна частина молекули) і залишок фосфорної кислоти (полярна гідрофільна «головка» молекули). Крім фосфоліпідів, важливим компонентом мембран є гліколіпіди й холестерин, роль якого особливо велика в плазматичних мембранах. Гліколіпіди нарівні з білками входять до складу іонних каналів, вони є рецепторами деяких класів речовин, що зумовлюють імунологічні властивості клітин, приймають участь у взаємодіях клітин.
Білки мембран, як правило, нерозчинні у воді, їх поділяють на 3 групи: структурні, ферментні й рецепторні (що специфічно зв’язують певні речовини). Білково-ліпідні комплекси є основою структури мембран та їх функціональної організації.
Вуглеводи присутні в основному у вигляді гліколіпідів, глікопротеідів і мукополісахаридів. Їх доля складає 2-10% сухої маси плазматичних мембран. У клітинах органоїдів їх може бути значно більше.
Доля води в мембранах може складати 20-35% маси. Мембрани, товщина яких становить 7-8 нм, можна побачити на електронно-мікроскпічних знімках, де вони виглядають трьохшаровими структурами. Мембрани різних клітин і різних органоїдів розрізняються між собою і структурою, і функціонально.
Згідно рідинно-мозаїчої моделі (рис. 1), основу мембрани складає бішар ліпідів, в якому неполярні "хвости" молекул обох шарів звернені один до одного, а гідрофільні "головки" – до оточуючого водяного середовища. Білки, у залежності від їх будови й функцій, розміщуються в мембрані по-різному. Деякі є периферичними, вони адсорбовані на поверхні мембран за рахунок електростатичних сил взаємодії з білками, що занурені в бішар повністю або частково (гідрофільними частинами), і називаються інтегральними. Інші інтегральні білки можуть пронизувати бішар повністю, зберігаючи виступаючі частини, які здатні взаємодіяти з молекулами або іонами оточуючого середовища. До числа інтегральних білків належать білки-переносники, що забезпечують транспорт речовин через мембрану, білки, що вистилають пори й формують іонні канали, деякі ферментні білки, тощо. Форма білкових молекул також може бути різною – від глобулярної до фібрилярної. До складу окремих білкових глобул може входити декілька поліпептидних ланцюжків.
Вуглеводні речовини розміщуються як на поверхні мембран (наприклад, мукополісахариди клітинної стінки), так і входять до складу мембран.
І білки, і ліпіди мембран не є жорстко закріпленими в певних позиціях. Зміна температури, електричного поля, хімічного оточення приводять до змін у структурі білків – конформаційних змін. Крім цього, відбувається обмін молекул як у середині кожного шару мембрани, так і між її шарами. Швидкість цього обміну також не змінюється. Усе це зумовлює велику чутливість мембрани до умов, в яких вона знаходиться.
Така схема будови мембрани добре узгоджується з більшістю експериментальних даних.
В окремих випадках експериментальні дані й рентгеноструктурний аналіз підтверджують справедливість глобулярної моделі. Один з її варіантів – ліпідні глобули, оточені білками, які забезпечують структурну стабільність мембрани – нині спростовується експериментальними даними. Другий варіант – білкові глобули, розміщені двома шарами, а порожнини між ними заповнені фосфоліпідами, полярні «головки» яких звернені в бік оточуючого середовища. Така структура, можливо, реалізована в мітохондріальних мембранах.
Біологічні мембрани виконують цілий ряд функцій.
Структурна функція. Зовнішня мембрана клітини відділяє її вміст від оточуючого середовища, визначає її форму. Оболонки органоїдів розділяють вміст різних зон клітини, забезпечують їх різний склад і, відповідно, різні біохімічні реакції в них.
Захисна функція. Якщо цитоплазматична мембрана захищає вміст клітин від небажаних впливів оточуючого середовища, то мембрани органоїдів виконують аналогічну функцію у відношенні до власного вмісту. Наприклад, мембрани лізосом захищають клітину від її ж літичних ферментів, яких достатньо, щоб викликати повний аутоліз (саморозпадання) клітин.
Транспортна функція. Здійснюється вона за рахунок наявності пор, каналів і білків-переносників. Транспорт речовин по цих структурах здійснюється вибірково й залежить від виду та складу речовин. Пори й канали мають малий діаметр (менше 1 нм), вислані білками, полярні групування яких взаємодіють з проникаючими в пори речовинами, сприяють або, навпаки, перешкоджають їх перенесенню. Гідрофобні групи взаємодіють з ліпідами мембрани. Проникливість мембрани може змінюватися в залежності від стану клітини.
На мембранах зосереджена більша частина ферментів клітини. Цим мембрани організовують біохімічні реакції, у тому числі й багатоетапні. Вони забезпечують суворо визначену направленість реакцій за рахунок визначеної послідовності в розміщенні ферментів. Яскравим прикладом є дихальні ланцюжки мітохондрій.
Електрогенна функція. Полягає в тому, що зовнішня мембрана клітин завдяки наявності АТФ-аз – переносників іонів створює різницю концентрацій K+, Na+, Ca+ і Cl– між цитоплазмою та міжклітинним середовищем. Різна проникливість мембрани для цих та інших іонів призводить до появи надлишкового від’ємного заряду в середині клітини за рахунок дифузії іонів К+ в оточуюче середовище (і, відповідно, різниці потенціалів між внутрішньою та зовнішньою сторонами мембрани). АТФ-аза (аденозинтрифосфатаза) – загальна назва білків, що розкладають АТФ на АДФ і фосфат. Внаслідок цього виділяється енергія, яка використовується для здійснення ендотермічних реакцій.
Мембрани створюють адгезію – зчеплення клітин. Ця властивість зумовлює існування багатоклітинних організмів. Адгезія відбувається, незважаючи на наявність одноіменного заряду клітин.
Основні механізми адгезії:
а) механічна взаємодія – зчеплення виступаючих ділянок мембран;
б) "склеювання" мембран з участю органічних солей кальцію, які взаємодіють з карбоксильними групами білків і фосфатними групами ліпідів;
в) взаємодія білків, що покривають мембрану, утворення пептидних зв’язків.
Мембрани виконують рецепторну функцію. На мембранах знаходяться рецепторні білки чи комплекси, які сприймають вплив оточуючого середовища (наприклад, взаємодіють з гормонами чи медіаторами) і, відповідно, міняють метаболізм клітини.
Антигенна функція. Полягає в тому, що на клітинній мембрані знаходяться білкові структури, що характерні для клітин даної тканини організму. Ці структури призначені для взаємного «пізнавання» клітин. Завдяки їх наявності можливе здійснення імунного захисту організму.
Матеріали для самоконтролю.
А. Завдання для самоконтролю (Тестові завдання)
Товщина біологічної мембрани:
10 А
10 нм
0,1 мкм
10 мкм
Рідко-мозаїчна модель біологічної мембрани включає в себе:
білковий шар, полісахариди і поверхневі ліпіди;
ліпідний моно шар і холестерин;
ліпідний біошар, білки, мікрофіламенти;
ліпідний біошар.
Товщина біологічної мембрани складає порядку:
0,01 нм;
0,1 нм;
10 нм;
100нм;
1 мкм.
Згідно рідко-мозаїчної моделі, біологічна мембрана складається:
з ліпідного шару;
двох шарів ліпідів з білковим шаром між ними;
двох шарів ліпідів, оточених зверху і знизу двома суцільними білковими шарами;
біліпідного шару, білків і мікроелементів;
шару ліпідів із вкрапленнями білків і вуглеводів.
Латеральною дифузією називається дифузія:
молекул з одного ліпідного шару в інший;
молекул через біологічну мембрану;
молекул у мембрані у межах одного шару;
білкових молекул із одного ліпідного шару в інший;
іонів через двошарову мембрану.
Перехід молекули з одного ліпідного шару в інший називається:
«фліп-флоп»-переходом;
полегшеною дифузією;
активним транспортом;
латеральною дифузією;
пасивним транспортом.
Ліпосомами називаються:
мономолекулярні шари на межі розділу гідрофобної і гідрофільної фази;
плоскі бішарові ліпідні мембрани;
біліпідні замкнуті структури;
шари ліпідів і білків, нанесені на поверхню води;
те ж саме, що і міцели.
Ліпіди в складі біологічних мембран знаходяться:
у твердому аморфному стані;
твердокристалічному стані;
рідкому аморфному стані;
рідкокристалічному стані;
правильної відповіді немає.
При фазовому переході мембран із рідкокристалічного в гель-стан площа мембрани, що приходить на одну молекулу ліпіду:
зменшується;
збільшується;
не змінюється.
При фазовому переході мембран із рідкокристалічного в гель-стан товщина мембрани:
зменшується;
збільшується;
не змінюється.
Б. Задачі для самоконтролю
Знайти коефіцієнт дифузії молекули сечовини якщо за 1 год вона продифундувала у воді на 3,1 мм.
Визначте рівноважний потенціал для іонів хлору в еритроцитах людини при температурі
 .
Концентрація іонів хлору всередині
клітини 80ммоль/л,
а ззовні – 3 ммоль/л.
.
Концентрація іонів хлору всередині
клітини 80ммоль/л,
а ззовні – 3 ммоль/л.Мембрана мітохондрії товщиною до 6 нм підтримує різницю потенціалів . Знайти напруженість електричного поля в мембрані.
Коефіцієнт дифузії глюкози
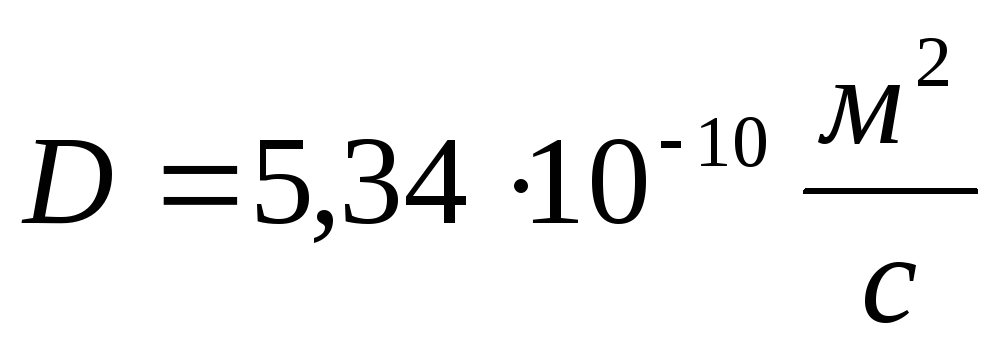 .
За який час молекула глюкози продифундує
на: а) 2мкм;
б) 0,4 м?
.
За який час молекула глюкози продифундує
на: а) 2мкм;
б) 0,4 м?
Література
Основна література:
Медична і біологічна фізика/ За ред. О. В. Чалого, 2-ге видання.- К.: Книга-плюс, 2005.
Медична і біологічна фізика / За ред. О. В. Чалого. Т. 1 К.: Віпол,1999; Т. 2- К.: Віпол, 2001.
Медична і біологічна фізика (практикум) за ред. О. В. Чалого. - К.: Книга-плюс, 2003.
Свердан П. Л. Вища математика: Аналіз інформації у математиці та медицині. -Львів: Світ, 1998.
Чалий О. В., Стучинська Н. В., Меленевська А. В. Вища математика. - К.: Техніка, 2001.
Костюк П. Г., Зима В. Л., Магура І. С., Мірошниченко М. С., Шуба М.Ф. Біофізика. – К.: Обереги, 2001.
Тиманюк В. А., Животова Е. Н. Биофизика. – Харьков, Изд-во НФАУ, 2003.
Зима В. Л. Біофизика. Збірник задач. - К.: В. шк.,2001.
Русяев В. Ф., Мищенко С. В., Пронина Н. В. Медицинская физика (сборник вопросов и задач). – Полтава, АСМИ 2001.
Іщейкіна Ю.О., Макаренко В.І., Тронь Н.В. Медична і біологічна фізика [Навчальний посібник] – Полтава: Шевченко Р.В., 2012. – 352 с., іл.
Додаткова література:
1. Ремизов А. Н. Медицинская и биологическая физика. – М.: Высш. шк., 1992.
2. Антонов В. Ф. и др. Биофизика. – М.: Владос, 2000.
3. Эссаулова И. Л., Блохина М. Е., Гонцов Л. Д. Руководство к лабораторным работам по медицинской и биологической физике. – М: Высш. шк., 1987.
4. Ремизов А. Н., Исакова Н. Х., Максина Л. Г. Сборник задач по медицинской и биологической физике. – М: Высш. шк., 1978.
5. Владимиров Ю. А., Рощупкин Д. И., Потапенко А. Я., Деев А, И . Биофизика. - М.: Медицина, 1983.
6. Рубин А. Б. Биофизика. – М.: Высш. шк., 1987.
7. Волькеннштейн М. В. Биофизика. – Высш. шк., 1987.
8. Самойлов О. В. Медицинская биофизика. – Л.: Изд-во ВМА, 1986.
9. Губанов Н. И., Утенбергов А. А. Медицинская биофизика. – М.: Медицина, 1981.
10. Лабораторный и лекционный эксперимент по медицинской и биологической физике / Под ред. Кройтора Д. С., Ремизова А. Н., Самойлова В. О. – Кишинев: Лумина, 1983.
11. Агапов Б. Т.,Максютин Г. В., Островерхов П. И. Лабораторный практикум по физике. – М.: Высш. шк., 1982.
12. Хакен Г. Синергетика. – М.: Мир, 1980.
13. Чернавский Д. С. Синергетика и информатика. – М.: УРСС, 2004.
14. Чалый А. В.,Цехмистер Я. В. Флуктуационные модели процессов самоорганизациии. К.: Випол, Випол, 19994.
15. Чалый А. В. Неравновесные процессы в физике биологии. – К.: Наук. думка, 1997.
16. Чалий О. В. Синергетичні принципи освіти та науки. – К.: Віпол, 2000.
Міністерство охорони здоров’я України
Вищий державний навчальний заклад України
"Українська медична стоматологічна академія"
«Затверджено»
на засіданні кафедри
медичної інформатики,
медичної і біологічної фізики
Завідувачка кафедри
професор__________ Іщейкіна Ю.О.
«27» серпня 2014 р.
Методичні вказівки
для самостійної роботи студентів
під час підготовки до практичного (семінарського) заняття
-
Навчальна дисципліна
Медична і біологічна фізика
Модуль №2
Основи біологічної фізики
Змістовний модуль
Біофізика мембранних процесів
Тема заняття
Мембранні потенціали спокою та дії.
Курс
1
Факультет
Медичний № 1, 2
Полтава – 2014
Актуальність теми.
Процеси життєдіяльності організмів супроводжуються появою в клітинах і тканинах електрорушійних сил. В нормально функціонуючій живій клітині, яка знаходиться в стані спокою, завжди існує різниця потенціалів між цитоплазмою і навколишнім середовищем. Нормальне функціонування організму неможливе без обміну інформацією між клітинами, одним зі способів якого є можливість генерації, передачі і сприйняття клітинами нервового імпульсу. Генерація і поширення електричних потенціалів — найважливіше фізичне явище в живих клітинах і тканинах, що лежить в основі збудливості клітин і проведенні збудження по клітинах, регуляції внутрішньоклітинних процесів, роботи нервової системи, регуляції м'язового скорочення. В організмі існують так звані збудливі клітини, до яких відносяться м'язові, нервові і секреторні. Так, м'язові клітини скорочуються, секреторні виділяють біологічно активні речовини, а нервові генерують електричні коливання — нервовий імпульс.
Завдяки
безпосередньому зв'язку біопотенціалів
з метаболічними процесами і фізіологічним
станом клітин, вони є показником різних
змін у клітинах у нормі і при патології.
Для більш ефективного і плідного
використання електрофізіологічних
методів у медицині є необхідним з'ясування
механізмів виникнення біоелектричних
потенціалів.Порушення
електричних
характеристик окремих клітин, нервових
волокон і цілих тканин, наприклад
серцевої тканини, призводить
до ряду серйозних захворювань. Тому
вивчення мембранних потенціалів є
важливим як для діагностики так і для
терапевтичних цілей.
Конкретні цілі.
Ознайомитись з методами електрофізіологічних досліджень мембранних потенціалів та застосуванням їх результатів в медицині.
Вивчити механізм виникнення потенціалів спокою та дії, а також процес поширення потенціалу дії в мієлінізованих та немієлінізованих нервових волокнах.
Засвоїти поняття: трансмембранний потенціал; потенціал спокою; рівноважний потенціал; потенціал Нернста; дифузний потенціал; потенціал Доннана; стаціонарний потенціал; потенціал дії або спайк; деполяризація; реполяризація; гіперполяризація; реверсія; воротні струми; стала довжини волокна.
Знати: рівняння для визначення рівноважних потенціалів (Нернста; Доннана; дифузійного; стаціонарного); феноменологічні рівняння Ходжкіна-Хакслі; телеграфне рівняння.
Оволодіти методикою розв’язування якісних і кількісних задач на визначення потенціалів спокою та дії за допомогою основних рівнянь.
Вміти застосовувати отримані знання при розв’язанні не стандартних задач практичного змісту.
Базові знання, вміння, навички, необхідні для вивчення теми (міждисциплінарна інтеграція)
|
Назви попередніх дисциплін |
Отримані навики |
|
1.Попередні (забезпечуючі дисципліни): Фізика, математика, хімія, біологія. |
Явище: поляризації. Поняття: електричний струм, аніон, катіон, аксоплазма, швановська клітина, перехват Ранв’є. Величини: електроємність, провідність, потенціал, концентрація іонів, потік іонів. Закон Ома, основне рівняння електродифузії, Теорію диф. рівнянь. Будову: мембранних насосів, цитоплазматичної мембрани, аксона, нервових волокон. Аналізувати фізичні процеси. Розв’язувати якісні і кількісні задачі. |
Завдання для самостійної роботи під час підготовки до заняття
Перелік основних термінів, параметрів, зарактеристик, які повинен засвоїти студент при підготовці до заняття.
Потенціал спокою – різниця потенціалів між цитоплазмою і навколишнім середовищем у нормально функціонуючій не збудженій клітині, що зумовлена відміною у концентраціях іонів по обидва боки мембрани.
У стані
спокою електричний потенціал цитоплазми
![]() є негативним відносно електричного
потенціалу зовнішнього середовища
клітини
є негативним відносно електричного
потенціалу зовнішнього середовища
клітини![]() .
Мембранний потенціал(потенціал
спокою)—
негативна величина, що дорівнює
.
Мембранний потенціал(потенціал
спокою)—
негативна величина, що дорівнює
![]() і становить у різних клітинах -30…-100мВ.
Так, потенціал спокою мієлінізованого
нервового волокна дорівнює -70мВ.
і становить у різних клітинах -30…-100мВ.
Так, потенціал спокою мієлінізованого
нервового волокна дорівнює -70мВ.
Розглядаючи різні умови руху іонів у просторі, можна одержати різні типи електричних потенціалів.
Рівноважний потенціал Нернста виникає при нерівномірному розподілі концентрації певних іонів у разі, коли мембрана проникна лише для цих іонів, і описується такою формулою:
![]() ,
,
де
се
і сі
— концентрація іонів відповідно зовні
та всередині клітини;
![]() — універсальна газова стала;Т
—
абсолютна температура;
— універсальна газова стала;Т
—
абсолютна температура;
![]() — число Фарадея,
z
– валентність іона.
— число Фарадея,
z
– валентність іона.
Дифузійний
рівноважний потенціал
встановлюється
в
середовищі
при наявності градієнта концентрацій
іонів різного
знака, що мають різну рухливість (![]() ),відповідно
до
формули:
),відповідно
до
формули:
![]()
Доннанівський потенціал виникає на мембрані, яка проникна для малих іонів різних знаків (Nа+, К+, Сl- та ін.) та непроникна для великих заряджених молекул (наприклад, білків, нуклеїнових кислот і інших великих органічних іонів), розташованих всередині клітинного простору. Найбільш характерна рівновага Доннана для мертвих клітин або для клітин з послабленим метаболізмом. Формула для потенціалу Доннана має такий вигляд:
![]()
де
![]() - концентрація білкових аніонів всередині
клітини,се
- концентрація неорганічних іонів
зовні.
- концентрація білкових аніонів всередині
клітини,се
- концентрація неорганічних іонів
зовні.
Стаціонарний
потенціал Гольдмана-Ходжкіиа-Катца
встановлюється на мембрані спричинений
не умовою рівноваги (![]() ),а
умовою стаціонарності, яка означає, що
повний потік, зумовлений потоками всіх
іонів, дорівнює нулю (
),а
умовою стаціонарності, яка означає, що
повний потік, зумовлений потоками всіх
іонів, дорівнює нулю (![]() ),
при цьому кожний з потоків у загальному
випадку відмінний від нуля (
),
при цьому кожний з потоків у загальному
випадку відмінний від нуля (![]() ).
).
У теорії стаціонарного потенціалу, розвиненої Д.Гольдманом, А.Ходжкіним і Б.Катцем, враховувалися внески в сумарний потік лише одновалентних іонів натрію, калію і хлору.
Розраховуючи мембранний потенціал, що виникає за цих умов, автори виходили з таких положень:
здійснюється умова стаціонарності (у стаціонарному стані сума потоків іонів через мембрану дорівнює нулю, хоча потоки окремих іонів можуть бути і відмінні від нуля):
![]()
у мембрані спостерігається сталість градієнта електричного потенціалу:
![]()
3) потік кожного сорту іонів підпорядковується електродифузному рівнянню Нернста-Планка:
![]() ,
,
де і – індекс сорту іонів (K+, Na+ і Cl-).
Інтегрування
рівнянь з урахуванням умови стаціонарності
![]() дає змогу одержати такий вираз для
мембранного потенціалу:
дає змогу одержати такий вираз для
мембранного потенціалу:

де
![]() - коефіцієнти проникності мембрани для
відповіднихіонів.
- коефіцієнти проникності мембрани для
відповіднихіонів.
У стані спокою співвідношення коефіцієнтів проникності мембрани аксона кальмара для іонів K+, Na+ і Cl- відносяться як 1:0,04:0,45.Так як в спокої проникність клітини для іонів калію набагато більше її проникності для інших іонів, то потенціал спокою визначається переважно різницею концентрацій іонів калію.
Підтримка різниці концентрацій іонів здійснюється за допомогою роботи іонних насосів, що використовують енергію АТФ. Na+,K+-насос, крім того, сприяє збільшенню трансмембранного потенціалу, тому що виводить три іони натрію і накачує в клітку тільки два іони калію, що збільшує позитивний заряд міжклітинного середовища, а отже, збільшує потенціал спокою.
Збільшення трансмембранної різниці потенціалів називається гіперполяризацією, зменшення — деполяризацією. В останньому випадку може навіть змінитися знак внутрішньоклітинного потенціалу.
Порушення роботи АТФ-азы, яке можна викликати, наприклад, дією серцевого глікозиду уабаїну, зменшує потенціал спокою. До такого ж ефекту приводить дія різної отрути (наприклад батрахотоксину), що збільшують проникність клітинної мембрани для іонів натрію. Деполяризацію викликає також гальмування процесів, що забезпечують у клітині синтез АТФ.
Внаслідок дії на клітину якого-небудь подразника її трансмембранний потенціал змінюється, виникає так званий потенціал дії, або спайк.
Потенціал дії – різниця потенціалів, що виникає при збудженні клітини і зумовлена зміною проникності мембрани для іонів.
Причиною такого коливання потенціалу спокою є зміна проникності мембрани для натрію, що, у свою чергу, викликано відкриттям натрієвих іонних каналів. Під час збудження проникності мембрани для іонів калію і натрію відносяться як 1:20. В результаті потік іонів натрію в клітину починає перевищувати потік іонів калію з клітини (мал.). Якщо раніше потенціал на мембрані був близький до рівноважного калієвого потенціалу, то тепер він наближається до рівноважного натрієвого, але не досягає його тільки внаслідок того, що провідності для іонів калію і хлору відмінні від нуля. В аксоні кальмара весь спайк триває близько 6мс.
Згідно гіпотези виникнення ПД у клітині відбувається деполяризація мембрани: негативний потенціал клітини наближається до нуля, а потім і зовсім змінює знак на протилежний (мал.). Останній процес називається реверсією мембранного потенціалу. Максимальне значення потенціалу дії складає за звичай 30...40мВ. Збільшення натрієвої провідності триває частки мілісекунд. Далі вона починає знижуватися, а калієва — зростати, у результаті чого в клітині відновлюється потенціал спокою. Цей процес називається реполяризацією клітинної мембрани.
Для відновлення потенціалу спокою клітині потрібний якийсь час. У процесі реполярнзації спочатку відбувається швидке наближення до нормального значення потенціалу спокою, потім швидкість зміни потенціалу зменшується, і клітина якийсь час перебуває в деполяризованому стані. Це називається слідовою деполяризацією. У деяких клітинах, навпаки, виникає слідова гіперполяризація, тобто в процесі реполярнзації різниця потенціалів починає перевищувати звичайну. Обоє ці відхилення від нормального значення потенціалу спокою називаються слідовими потенціалами.
Іони
Na+
входячи всередину клітини, призводять
до ще більшого зростання потенціалу
![]() на внутрішньому боці мембрани, що
викликає, в свою чергу, подальше зростання
проникності мембрани для іонів Na+
і, як наслідок, зростання швидкості їх
проникнення всередину клітини. Зростання
потенціалу на внутрішньому боці мембрани
викличе, з деяким запізненням, повільне
збільшення проникності мембрани для
іонів K+,
внаслідок пасивної дифузії вони почнуть
виходити назовні. При цьому спостерігається
повернення мембранного потенціалу
на внутрішньому боці мембрани, що
викликає, в свою чергу, подальше зростання
проникності мембрани для іонів Na+
і, як наслідок, зростання швидкості їх
проникнення всередину клітини. Зростання
потенціалу на внутрішньому боці мембрани
викличе, з деяким запізненням, повільне
збільшення проникності мембрани для
іонів K+,
внаслідок пасивної дифузії вони почнуть
виходити назовні. При цьому спостерігається
повернення мембранного потенціалу
![]() від максимального до вихідного значення
(від +40 до -70 мВ). Це процес так званої
реполяризації (див. рис.).
від максимального до вихідного значення
(від +40 до -70 мВ). Це процес так званої
реполяризації (див. рис.).
Іони Na+, що увійшли у клітину під час зростання ПД, можуть вийти назовні тільки внаслідок роботи Na+,K+-насоса. Після ПД спостерігається період не збудження або абсолютний рефрактерний період, під час якого не може виникнути наступний ПД. Він триває від 0.5 до 2мс. За цей час внаслідок роботи Na+,K+-насоса відбувається відновлення вихідних значень концентрацій іонів Na+ і K+.
Таким чином, формування ПД спричинене двома іонними потоками крізь мембрану: потік іонів Na+ всередину клітини викликає перезарядку мембрани, а протилежно спрямований потік K+ - відновлення вихідного значення потенціалу спокою. Потоки зсунуті у часі, завдяки чому можлива поява ПД (рис.).
Згідно з моделлю Ходжкіна і Хакслі зміна іонних провідностей при зсувах мембранних потенціалів спричинена впливом електричного поля на просторовий розподіл у мембрані заряджених активуючих частинок.
На рис.
подано еквівалентну
схему
елемента збудливої мембрани нервового
волокна. Вона становить електричне коло
з чотирма паралельними вітками. Одна з
них містить електричну ємність, а інші
відтворюють натрієву (![]() ),
калієву (
),
калієву (![]() )
провідності мембрани і так звану
провідність відтоку (
)
провідності мембрани і так звану
провідність відтоку (![]() ).
У кожну з цих віток включені джерела
електрорушійної сили (
).
У кожну з цих віток включені джерела
електрорушійної сили (![]() ),
які дорівнюють відповідним рівноважним
потенціалам Нернста:
),
які дорівнюють відповідним рівноважним
потенціалам Нернста:
![]() ;
;
![]() ;
;![]() .
.
Вираз для електричного струму крізь мембрану можна подати у такому вигляді:
![]()
![]() .
.
Перший доданок у рівнянні - це ємнісний струм, пов'язаний зі зміною різниці потенціалів на мембрані, а другий доданок - іонний струм.
Експериментально
Ходжкін і Хакслі довели, що іонні струми
![]() прямо пропорційні різниці потенціалів
на мембрані
прямо пропорційні різниці потенціалів
на мембрані![]() і рівноважному мембранному потенціалу
Нернста для відповідного сорту іонів:
і рівноважному мембранному потенціалу
Нернста для відповідного сорту іонів:
![]() ;
;
![]()
![]() .
.
Останнє рівняння в враховує іонні струми всіх інших іонів, а саме: Сl- , Са2+ тощо. Коефіцієнти gi, в рівняннях характеризують провідності каналів. Провідність каналів може змінюватись внаслідок їх активації.
Один із постулатів математичної моделі мембрани говориться проте, що у внутрішній структурі мембрани існують деякі зарядженні частинки, які керують провідністю каналів. В залежності від величини напруженості прикладеного електричного поля ці гіпотетичні частинки можуть переміщуватися в мембрані, і тим самим збільшувати або зменшувати потоки іонів натрію і калію крізь канали.
Потенціалкеровані натрієві, калієві та кальцієві канали відповідають за генерацію поширюваних електричних сигналів у нейронах та інших збудливих клітинах (збудливим клітинам властива можливість генерувати потенціали дії). Ці канали також функціонують у незбудливих клітинах, зокрема клітинах глії, лімфоцитах тощо. Потенціалкеровані іонні канали реагують на зміни напруженості електричного поля в мембрані конформаційними змінами білків, що утворюють іонні канали. Сучасні дослідження показали, що перехід від конформації, що відповідає закритому стану, до конформації відкритого каналу (активація) зумовлений транслокацією через мембрану рухливих зарядів, еквівалентних приблизно 10 елементарним зарядам. Це переміщення зарядів може бути безпосередньо виміряне як воротний струм.
Воротний струм являє собою компонент струму зміщення, тобто ємнісного струму. Його реєстрація має певні труднощі. При активації іонного каналу внаслідок впливу електричного поля на сенсор напруги в мембрані переміщається лише близько 10 елементарних зарядів, тоді як іонний струм, що виникає при цьому, зумовлений проходженням через канав тисяч іонів за одну мілісекунду. Для виділення воротного струму необхідно знизити до мінімуму іонні струми, що протікають через мембрану. Тому проникні через мембрану іони замінюють на непроникні. Використовують також речовини, які блокують іонні струми, але не впливають на воротні механізми. Незважаючи на ці заходи, струм витоку зберігається.
Воротні струми натрієвих каналів уперше було зареєстровано на перфузованому внутрішньоклітинному гігантському аксоні кальмара К.Армстронгом і Ф.Безаніллою, а також Р.Кейнсом і Е.Рохасом. Для пригнічення натрієвого струму використовували тетродотоксин. Крім того, іони натрію в зовнішньому розчині замінювали великими непроникними катіонами, а всередину аксона замість іонів калію вводили іони цезію, що блокують калієві канали. Для усунення струму витоку та виділення воротного струму здійснювали багаторазові симетричні позитивні й негативні зсуви мембранного потенціалу. Струми, які реєструвалися и алгебраїчно підсумовували. При цьому було виділено асиметричний струм зміщення, що був ідентифікований як воротний струм натрієвих каналів
Фізична інтерпретація моделі Ходжкіна-Хакслі вимагала наявності всередині мембрани деяких заряджених частинок, які б мали переміщатися в залежності від зовнішнього електричного поля. Таким чином, для підтвердження другого постулату моделі необхідно було зареєструвати переміщення заряджених частинок всередині мембрани при зміні мембранного потенціалу, тобто зареєструвати так звані воротні струми. Складність виявлення воротних струмів полягала в тому, що активуючих частинок всередині мембрани дуже мало и, тому, мале значення воротного струму в порівнянні з іонними струмами, які проходять через мембрану.
Для виявлення воротних струмів за допомогою блокаторів ТТХ и ТЕА, а також заміною іонів натрію в навколишньому розчині на іони трису, виключали іонні струми; потім ступінчато змінювали напругу на мембрані и реєстрували появу воротного струму натрієвого каналу, який виявився в 103 раз слабшим натрієвого струму.
Зміна в часі воротного струму в аксоні кальмара було взаємопов’язана зі зміною натрієвого струму. Таким чином на досліді було доведено існування воротних струмів.
Потенціал дії, що виник на певній ділянці нервової клітини, швидко розповсюджується вздовж її поверхні завдяки локальним струмам між збудженими і не збудженими ділянками нервового волокна (рис.). Локальні струми чинять подразнюючу дію на сусідні не збуджені ділянки і викликають зміну проникності мембрани. Локальні струми деполяризують мембрану до певного рівня, а ПД на кожній ділянці виникає внаслідок іонних потоків Nа+ та К+, спрямованих перпендикулярно напрямку поширення збудження.
Нервові волокна поділяються на миєлінізовані (м’якотні) і немієлінізовані (безм’якотні). Мієлінізоване нервове волокно складається з осьового циліндра, покритого цитоплазматичною мембраною, утримуючою аксоплазму (мал.). Довкола нього багаторазово обертаються швановські клітини (у периферичній нервовій системі) чи олігодендроглії (у центральній нервовій системі), шари яких зливаються й утворять мієлінізовану оболонку нервового волокна. Через рівні проміжки (від 0,2 до 2мм), характерні для даної клітини, ця оболонка переривається, і мембрана осьового циліндра залишається відкритою. Такі ділянки волокна називаються перехопленнями Ранвье. Їхня довжина складає приблизно 1мкм. Мієлінова оболонка, що складається з мембранних ліпідів і білків, є надійним ізолятором нервової клітки, завдяки їй збудження може виникнути тільки на оголеній ділянці мембрани аксона. Немієлінізовані нервові волокна не мають такої щільної жирової оболонки. Швановска клітина оточує їх тільки один раз.
Порушення якої-небудь ділянки немієлінізованого нервового волокна призводить до локальної деполяризації мембрани. У той же час інша (не збуджена) частина мембрани зберігає свою звичайну різницю потенціалів: зовнішнє середовище заряджене позитивно, а внутрішня — негативно. Між збудженою і не збудженою областями виникають місцеві струми (мал.). Це призводить до деполяризації сусідньої ділянки, що, у свою чергу, деполяризує наступний. Приєднавши електроди осцилографа до двох ділянок клітинної мембрани, можна прослідкувати поширення потенціалу дії. Такий спосіб проведення збудження називається неперервним.
У мієлінізованих нервових волокнах неперервне проведення нервового імпульсу неможливе. Збудження (деполяризація) може виникати не по всій довжині мембрани, а тільки в перехватах Ранв’є. Деполяризація однієї такої ділянки А викликає деполяризацію сусідньої ділянки Б (мал.). Далі збудження здатне перейти тільки до ділянки В, тому що А на протязі деякого часу залишається нечутливим до збудження (рефрактерним). По цій причині імпульс поширюється по нервовому волокну тільки в одному напрямку. Виникаючий потенціал дії в кілька разів перевищує поріг, необхідний для виникнення збудження в наступному перехваті Ранв’є, що, таким чином, щораз підсилює сигнал, що слабшає в результаті опору міжтканинної рідини й аксоплазми, і діє подібно ретранслятору. Механізм поширення збудження по мієлінізованих волокнах називається стрибкоподібним або сальтаторним.
Сальтаторний механізм вигідніше безперервного, тому що дозволяє збільшити швидкість проведення нервового імпульсу і є більш економічним з енергетичної точки зору: деполяризуются тільки невеликі ділянки мембрани, виникають менші втрати іонів, отже, клітині приходиться витрачати менше енергії для забезпечення роботи, Na+,K+-насосів.
При одному з автоімунних захворювань — розсіяному склерозі — імунна система організму руйнує мієліновую оболонку, відбувається оголення (демієлінізація) нервових волокон. При цьому проведення нервових імпульсів через уражену ділянку порушується, що призводить до різних проявів: порушенню зору і координації, м'язовій слабкості, підвищенню м'язового тонусу й ін. Демієлінізація може відбуватися і при деяких інших захворюваннях: невралгіях, радикулітах, різних поліневропатіях.
Розподіл
потенціалу дії
![]() в залежності від відстані
в залежності від відстані![]() і часу
і часу![]() по немієлінізованому нервовому волокні
визначається так званимтелеграфним
рівнянням:
по немієлінізованому нервовому волокні
визначається так званимтелеграфним
рівнянням:
![]()
де
![]() — діаметр волокна;
— діаметр волокна;![]() — товщина мембрани;
— товщина мембрани;![]() — електроємність;
— електроємність;![]() — питомий опір аксоплазми;
— питомий опір аксоплазми;![]() — питомий опір мембрани, що різко
знижується під час збудження.
— питомий опір мембрани, що різко
знижується під час збудження.
Розв’язок
цього рівняння в стаціонарному режимі
(при
![]() )
має наступний вид:
)
має наступний вид:
![]() , (12.5.2)
, (12.5.2)
де
![]() — потенціал у точці х=0;
— потенціал у точці х=0;![]() — постійна довжини волокна, яка дорівнює
— постійна довжини волокна, яка дорівнює
![]() , (12.5.3)
, (12.5.3)
При
віддаленні від точки прикладання
збудження на величину
![]() потенціал зменшується в е раз. Клітині
вигідніше мати великі значення
потенціал зменшується в е раз. Клітині
вигідніше мати великі значення![]() ,
тому що при цьому затухання імпульсу
відбувається повільніше. Відсутність
повного затухання імпульсу пояснюється
тим, що кожний наступний потенціал дії
підсилює сигнал.
,
тому що при цьому затухання імпульсу
відбувається повільніше. Відсутність
повного затухання імпульсу пояснюється
тим, що кожний наступний потенціал дії
підсилює сигнал.
Швидкість
проведення нервового імпульсу
по немієлінізованих нервових волокнах,
так само як і постійна довжини, пропорційна
квадратному кореню з діаметра волокна.
Збільшення діаметра сприяє збільшенню
![]() і швидкості поширення збудження. Цим
пояснюється існування гігантських
аксонів головоногих молюсків. Швидкість
проведення збудження по немієлінізованому
волокні діаметром 1мкм складає тільки
2м/с, тоді як для волокон діаметром
0,5—1мм ця величина вже досягає 20м/с.
і швидкості поширення збудження. Цим
пояснюється існування гігантських
аксонів головоногих молюсків. Швидкість
проведення збудження по немієлінізованому
волокні діаметром 1мкм складає тільки
2м/с, тоді як для волокон діаметром
0,5—1мм ця величина вже досягає 20м/с.
Для мієлінізованих нервових волокон швидкість проведення збудження залежить від довжини міжперехватних ділянок (ділянок між двома перехопленнями Ранв’є). Замічено, що час, необхідне для передачі сигналу від одного перехоплення до іншого, приблизно однаково і складає близько 0,07мс. У той же час довжина міжперехватних ділянок пропорційна діаметру волокна. Таким чином, швидкість проведення нервового імпульсу по м’якотних волокнах пропорційна їхньому діаметру. Тому в організмів, що мають мієлінізовані волокна, відпала необхідність значно збільшувати їхній діаметр. Таке волокно діаметром 20мкм проводить збудження зі швидкістю 120м/с.
Вимірювання швидкості проведення нервового імпульсу часто проводять у медичній практиці: при порушенні мієліновой оболонки швидкість поширення збудження знижується.
Для
нервових волокон приблизно однакового
діаметра швидкість поширення збудження
залежить від величини так званого
фактора надійності
![]() ,
що виражається в такий спосіб:
,
що виражається в такий спосіб:
![]() (12.5.4)
(12.5.4)
де
![]() — амплітуда потенціалу дії;
— амплітуда потенціалу дії;![]() — критичний рівень деполяризації, що
викликає генерацію потенціалу дії.
Звичайно фактор надійності складає
5—6, тобто потенціал дії здатний збудити
наступна ділянка мембрани, тому що його
величина значно перевищує граничний
рівень деполяризації. Дія місцевих
анестетиків (наприклад новокаїну)
зводиться до зниження величини
— критичний рівень деполяризації, що
викликає генерацію потенціалу дії.
Звичайно фактор надійності складає
5—6, тобто потенціал дії здатний збудити
наступна ділянка мембрани, тому що його
величина значно перевищує граничний
рівень деполяризації. Дія місцевих
анестетиків (наприклад новокаїну)
зводиться до зниження величини![]() і збільшенню
і збільшенню![]() за рахунок інактивації натрієвих
каналів, що приводить до зниження фактора
надійності.
за рахунок інактивації натрієвих
каналів, що приводить до зниження фактора
надійності.
Як відомо, швидкість поширення ПД залежить від того, чи має аксон мієлінову оболонку, чи така оболонка відсутня. У першому випадку розповсюдження нервового імпульсу відбувається стрибками через перехвати Ранв'є і тому має значно більшу швидкість, ніж у другому випадку.
Орієнтовна карта для самостійної роботи з літературою по темі.
|
№ |
Основні завдання |
Вказівки |
|
|
1 |
2 |
3 |
|
|
1 |
Вивчити основні поняття |
Дати означення таких понять: трансмембранний потенціал; потенціал спокою; рівноважний потенціал; потенціал Нернста; дифузний потенціал; потенціал Доннана; стаціонарний потенціал; потенціал дії або спайк; деполяризація; реполяризація; гіперполяризація; реверсія; воротні струми. (використати конспект лекцій) |
|
|
2 |
Вивчити основні характеристики |
Дати означення сталої довжини волокна. |
|
|
3 |
Засвоїти закономірності |
Записати: рівняння для визначення рівноважних потенціалів (Нернста; Доннана; дифузійного; стаціонарного); феноменологічні рівняння Ходжкіна-Хакслі. |
|
|
4 |
Ознайомитись із методами |
Ознайомитись з методами електрофізіологічних досліджень мембранних потенціалів та застосуванням їх результатів в медицині |
|
|
5 |
Вивчити механізм виникнення потенціалів. |
Вивчити механізм виникнення потенціалів спокою та дії, а також процес поширення потенціалу дії. |
|
4.2. Теоретичні питання до заняття
Що являє собою мембранний потенціал спокою?
Вкажіть, як співвідносяться між собою у стані спокою проникності мембран живих клітин для іонів.
Які умови враховуються при визначенні мембранного рівноважного потенціалу Нернста?
Вкажіть величини рівноважних потенціалів Нернста для іонів Na+, К+ і Сl- в нормі.
Які умови враховуються при визначенні рівноважного потенціалу Доннана?
Вкажіть, для яких випадків доннанівський потенціал ближче всього до потенціалу клітини. Які умови враховуються при визначенні дифузійного мембранного потенціалу?
Які умови враховуються при визначенні стаціонарного мембранного потенціалу?
Що таке потенціал дії?
Вкажіть причини утворення потенціалу дії.
Як співвідносяться в початковий момент збудження проникності мембран для іонів у живих клітинах?
Чим визначається форма початкової фази деполяризації мембрани при формуванні потенціалу дії?
Чим визначається фаза реполяризації мембрани при формуванні потенціалу дії? (α=ІІ)
Вкажіть тривалість потенціалу дії в нормі для аксону кальмара. (α=І)
Чи виникне потенціал дії, якщо з навколишнього середовища клітини видалити повністю іони натрію?
Які особливості поширення потенціалу дії по нервових волокнах?
Матеріали для самоконтролю.
А. Завдання для самоеревірки (тестові завдання)
Мембранний потенціал спокою…
а) ![]() ;
;
б) ![]() ;
;
в) ![]() .
.
Потенціал спокою визначається…
а) лише різною концентрацією іонів по різні сторони мембрани;
б) лише дифузією іонів крізь мембрану;
в) різною концентрацією іонів по різні сторони мембрани і дифузією іонів крізь мембрану;
г) дифузією нейтральних частинок крізь мембрану;
д) лише активним транспортом іонів.
У спокої потенціал нервової клітини наближається до рівноважного:
а) кальцієвого потенціалу;
б) натрієвого потенціалу;
в) хлорного потенціалу;
г) калієвого потенціалу;
д) потенціалу протонів.
Потенціал, який виникає на мембрані, яка проникна для малих іонів різних знаків (Nа+, К+, Сl- та ін.) та непроникна для великих заряджених молекул (наприклад, білків, нуклеїнових кислот і інших великих органічних іонів), розташованих всередині клітинного простору називається…
а) потенціалом спокою;
б) дифузним;
в) стаціонарним;
г) доннанівським;
д) потенціалом дії.
Різниця потенціалів, що виникає при збудженні клітини і зумовлена зміною проникності мембрани для іонів називається …
а) потенціалом спокою;
б) натрієвим потенціалом;
в) хлорним потенціалом;
г) калієвим потенціалом;
д) потенціалом дії.
Під час генерації потенціалу дії потенціал нервової клітки наближається до рівноважного:
а) кальцієвого потенціалу;
б) натрієвого потенціалу;
в) хлорного потенціалу;
г) калієвого потенціалу;
д) потенціалу протонів.
Внутрішньоклітинне середовище заряджене в порівнянні з позаклітинним:
а) у спокої — негативно, на максимумі потенціалу дії — позитивно;
б) у спокої — позитивно, на максимумі потенціалу дії — негативно;
в) завжди позитивно;
г) завжди негативно.
Проникність мембрани для іонів калію в спокої:
а) значно більша проникності для іонів натрію;
б) значно менше проникності для іонів натрію;
в) приблизно дорівнює проникності для іонів натрію.
Під час генерації потенціалу дії проникність мембрани для іонів калію:
а) більша проникності для іонів натрію;
б) менша проникності для іонів натрію;
в) приблизно дорівнює проникності для іонів натрію.
Збільшення трансмембранної різниці потенціалів називається …, зменшення — ….
а) деполяризацією; гіперполяризацією;
б) гіперполяризацією; деполяризацією;
в) реполяризацією; деполяризацією;
г) деполяризацією; реполяризацією;
д) рефрактерністю; реполяризацією.
Постійною довжини нервового волокна називається відстань, на якій:
а) швидкість поширення нервового імпульсу зменшується в е раз;
б) швидкість поширення нервового імпульсу збільшується в е раз;
в) швидкість поширення нервового імпульсу зменшується в 10 разів;
г) швидкість поширення нервового імпульсу збільшується в 10 разів;
д) потенціал зменшується в е раз.
Швидкість поширення нервового імпульсу пропорційний (D — діаметр волокна):
а)
![]() для немієлінованих і мієлінізованих
нервових волокон;
для немієлінованих і мієлінізованих
нервових волокон;
б) D для немієлінізованих і мієлінізованих нервових волокон;
в)
![]() для
немієлінізованих
і D
для мієлінізованих
нервових волокон;
для
немієлінізованих
і D
для мієлінізованих
нервових волокон;
г)
D
для немієлінізованих
і![]() для мієлінізованих
нервових волокон;
для мієлінізованих
нервових волокон;
д) D2 для немієлінізованих і мієлінізованих нервових волокон.
Б. Задачі для самоконтролю
Задача 1. Розрахуйте рівноважні мембранні потенціали для іонів натрію, калію і хлору для мембрани еритроциту, якщо їхня позаклітинна концентрація відповідно дорівнює: 155; 5; 4ммоль/л, а внутрішньоклітинна —19; 136; 120ммоль/л. Температура 37°С.
Відповідь: 57мВ; -89мВ; -92мВ.
Задача 2. Як відомо, концентрація іонів як Na+, так і Сl- зовні клітини перевищує їх концентрацію всередині приблизно на порядок. Рухливість цих іонів, знайдена в експерименті, становить b+=5.2*10-8м2/с*В; b-=7.9*10-8м2/с*В. Виконати чисельну оцінку дифузійного потенціалу, що спричинений різними рухливостями іонів, якщо T=293K.
Відповідь: 12мВ.
Задача 3. У внутрішньоклітинному середовищі присутні аніони білкового походження концентрація яких [Р-]=300мМ/л, кожна молекула містить п=1 заряд. Концентрація іонів зовні клітини се=4102мМ/л. Оцінити потенціал Донна на при температурі 20оС.
Відповідь: 0,9мВ.
Задача 4. У спокої проникності мембрани для іонів калію і натрію відносяться як РК:PNа=1:0,04, а при збудженні — Рк:PNa=1:20. Внутрішньоклітинна концентрація іонів калію складає [К+|=350ммоль/л, позаклітинна — у 50 разів менше; внутрішньоклітинна концентрація іонів натрію [Na+]і=50ммоль/л, позаклітинна — у 10 разів вище. Визначите рівноважний потенціал і для кожного з цих іонів, величину потенціалу спокою і потенціалу дії. Температура клітини 27°С.
Відповідь: -99мВ, 59мВ
Задача
5. Обчислити діаметр D
аксона,
якщо питомий опір одиниці
товщини мембрани
![]() =80мОм*м2,
питомий опір аксоплазми
=80мОм*м2,
питомий опір аксоплазми
![]() =0,4Ом*м,
постійна довжини
=0,4Ом*м,
постійна довжини![]() =4,5мм.
=4,5мм.
Відповідь: 0,4мм.