

Р ефлексы

 Местные
Истинные
Местные
Истинные
“Простые” Объединения безусловных
рефлексов



 Инстинкты
Условные рефлексы
Инстинкты
Условные рефлексы
Одиночные Динамический стереотип
поведения
В рефлексе как сложном нервном процессе различают четыре звена: 1) раздражение рецепторов, восприятие изменений внешней или внутренней среды и проведение нервных импульсов по афферентным путям в центральный отдел, 2) развертывание нервного процесса в центральном отделе, 3) проведение нервных импульсов по эфферентным путям, вызывающее или регулирующее функцию органа и 4) проведение афферентных импульсов из функционирующего органа в центральный отдел (обратная информация), что позволяет регулировать интенсивность и характер деятельности органа.
Таким образом, определяющим для формирования рефлекса является строение и локализация в нервной системе центрального отдела рефлекторной дуги или рефлекторного кольца. В том случае, если центральный отдел как таковой отсутствует или располагается вне ЦНС говорят о местных рефлексах, при его локализации в ЦНС - об истинных рефлексах.
Время от начала стимула до реакции эффектора называется временем рефлекса. В большинстве случаев оно определяется главным образом временем проведения в афферентных, эфферентных путях и в центральной части рефлекторной дуги. Сюда следует еще добавить а- время трансформации в рецепторе стимула в распространяющийся электрический имбульс, б- время передачи через синапсы (синаптическую задержку), в- время передачи от эфферентного пути к эффектору, г- время активации эффектора, путем возбуждения мембраны (например, электромеханическое сопряжение).
Любой рефлекторный акт всегда представляет собой результат взаимодействия различных рефлексов. Морфологической основой для такого взаимодействия являются две особенности строения нервной системы: наличие вставочных нейронов на которых могут сходиться несколько рефлекторных путей и превышение общего числа сенсорных (чувствительных) входов над числом двигательных выходов и нейронов.
Взаимодействие между рефлексами может осуществляться по типу усиления или ослабления вызываемой реакции. Рефлексы, усиливающие друг друга, называются аллиированными. Так, слюноотделительный рефлекс можно усилить раздражением тактильных и вкусовых рецепоров полости рта. Рефлексы препятствующие, тормозящие друг друга получили название антагонистических. Например, рефлекс глотания тормозит рефлекс вдоха. Это легко можно проверить, пытаясь проглотить слюну на фазе вдоха.
Взаимодействие рефлексов обеспечивается такими механизмами, как конвергенция, иррадиация, возбуждение, реципрокность возбуждения и торможения, доминанта, обратная связь, общность конечного пути.
МЕСТНЫЕ РЕФЛЕКСЫ. Эта группа рефлексов представлена аксон-рефлесом и узловыми (ганглионарными) рефлексами.
Аксон-рефлекс - (axon-reflexus) - общее название рефлексов, осуществляющихся по разветвлениям аксона без участия тела нейрона; рефлекторная дуга аксон-рефлекса не содержит синапсов и тел нейронов, возбуждение идет афферентно по одной ветви аксона, затем передается на другую ветвь и к эфферентному органу. В аксон-рефлексе практически отсутствует центральный отдел. С помощью аксон-рефлекса возможно переключение сигнала и с афферентного нейрона на эфферентный в периферическом ганглии (узле). Благодаря аксон-рефлексу может происходить регуляция внутренних органов и сосудов в определенной степени независимо от ЦНС.
Ганглионарные (узловые) рефлексы - В свое время А.С.Догель высказал предположение о том, что в вегетативных ганглиях, помимо эффекторных нейрогнов, имеются также и чувствительные нейроны. Это послужило основой для изучения вегетативных рефлексов, замыкающихся в вегетативных ганглиях. Ряд авторов считают, что при децентразизации звездчатого узла могут быть вызваны потовые и сосудистые рефлексы с передней конечности. Есть много наблюдений, свидетельствующих о том, что превертебральные ганглии могут явиться механизмом, определяющим рефлексы между различными частями кишечника. Однако, справедливости ради, следует отметить, что имеются исследователи отрицающие наличие таких рефлексов на основании утверждения о технической сложности полной децентрализации ганглиев.
ИСТИННЫЕ РЕФЛЕКСЫ характеризуются тем, что их центральное звено располагается в центральной нервной системе, т.е. они осуществляются с обязательным участием ЦНС.
По локализации центрального звена рефлексы делятся на спинальные, бульбарные, мезэнцефалические, мозжечковые, диэнцефалические, корковые.
Положив в основу классификации рефлексов анатомический принцип локализации центрального отдела рефлекса в ЦНС И.П.Павлов выделил среди истинных рефлексов
1. простые (спинномозговые)
2. усложненные (продолговатый мозг)
3. сложные (средний мозг)
4. сложнейшие (выше расположенные отделы головного мозга).
Приведенные выше классификации подразумевают локализацию центрального звена лишь на одном уровне.
Вместе с тем систематическими исследованиями Э.А.Асратяна и его сотрудников было убедительно показано, что центральная часть дуги безусловного рефлекса не однолинейна, проходит не через какой-нибудь один уровень мозга, а состоит из многих ветвей, каждая из которых проходит по основным отделам ЦНС - спинному мозгу, продолговатому мозгу и т.п. Самая верхняя часть дуги проходит через кору большого мозга, является корковым представительством данного безусловного рефлекса и олицетворяет кортиколизацию соответствующей функции организма. Наличие кортикального представительства у практически любого безусловного рефлекса создает анатомические предпосылки для формировани условных рефлексов первого порядка как синтеза двух или нескольких безусловных.
БЕЗУСЛОВНЫЕ РЕФЛЕКСЫ - врожденные, генетически детерминированные реакции, обусловленные жесткими нейронными связями в цепочке рецептор-эффектор. Характерной особенностью безусловных рефлексов является то, что их реализация определяется как внутренними детерминантами, так и внешней стимульной программой.
Примером простых безусловных рефлексов могут служить спинномозговые рефлексы. Рефлекторная деятельность спинного мозга обеспечивается сегментарными рефлекторными дугами. Такая дуга состоит из рецептивного поля, из которого импульсация по чувствительному волокну афферентного нейрона спинального ганглия, а затем по аксону этого же нейрона через задний корешок входит в спинной мозг. Далее возможно два варианта:
- аксон афферентного нейрона может идти прямо к мотонейрону переднего рога, аксон которого подходит к мышце (эфферентное звено). В результате формируется моносинаптическая (двухнейронная) рефлекторная дуга.
- аксон афферентного нейрона передает сигнал на вставочный нейрон, а последний на эфферентный - формируется полисинаптическая рефлекторная дуга.
По количеству центральных синапсов в рефлекторной дуге выделяют моносинаптические и полисинаптические рефлекторные дуги.
Моносинаптические рефлексы. Рефлекс растяжения, вызванный растяжением мышцы - сухожильный рефлекс. Кратковременное растяжение мышцы, вызываемое легким ударом молоточка по сухожилию вызывает после латентного периода сокращение той мышцы или даже части мышцы, которую раздражают. Афферентные волокна мышечных веретен образуют синапсы на гомонимных -мотонейронах (т.е. мотонейронах их собственных мышц). Залп потенциалов действия, поступающий в спинной мозг по афферентному нейрону вызывают моносинаптические возбуждающие постсинаптические потенциалы (ВПСП) в соответствующих гомонимных мотонейронах, причем некоторые ВПСП достигают надпорогового уровня и вызывают слабое сокращение мышцы. В эту группу рефлексов входят рефлекс закрывания рта при постукивании по подбородку, рефлексы на растяжение двуглавой мышцы постукиванием по ее сухожилию около локтя или на удар по лучевой кости, растяжение трехглавой мышцы плеча путем постукивания по ее сухожилию чуть выше локтевого отростка, растяжение четырехглавой мышцы постукиванием ниже коленной чашечки, растяжение трехглавой мышцы голени постукиванием по пяточному сухожилию. Рефлексы называют в зависимости от раздражаемого сухожилия, например, коленный (пателярный), ахиллов и т.п.
Однако выражение “сухожильный рефлекс” вводит в заблуждение, поскольку здесь рефлекс запускается путем активации первичных окончаний мышечных веретен при растяжении мышцы. В связи с чем используют термины рефлекс растяжения и миотатический рефлекс.
Выделяют две формы рефлекса на растяжение в зависимости от способа раздражения: тонический рефлекс на растяжение - возникает в ответ на длительное растяжение, продолжающееся десятки секунд и фазический рефлекс на растяжение - в ответ на очень кратковременное растяжение мышцы , например, при ударе молоточком.
Рефлексы растяжения можно класссифицировать по природе использованного раздражитетя. Так рефлексы, получаемые путем постукивания молоточком по сухожилию называют Т-рефлексами, а путем электрической стимуляции афферентных нервных волокон - Н-рефлексами (по имени P.Hoffmann).
После Т-рефлекса или Н-рефлекса тонус мышц резко падает на короткий период (100-500 мс). Этот пострефлекторный период молчания обусловлен сочетанием ряда факторов:
- синхронное рефлекторное сокращение снимает напряжение мышечных веретен и, следовательно, уменьшает или прекращает тонический возбуждающий афферентный вход от первичных окончаний мышечных веретен.
- рефлекторное сокращение активирует сухожильные органы Гольджи, которые обладают тормозным действием на соответствующие мотонейроны.
- синхронное возбуждение мотонейронов на короткое время усиливает торможение, производимое клетками Реншоу.
- гиперполяризующие следовые потенциалы после потенциалов действия в мотонейронах, участвующих в рефлексе, временно снижают их возбудимость.
Полисинаптические рефлексы. За исключением моносинаптического рефлекса растяжения, все рефлекторные дуги включают несколько последовательных центральных нейронов и образуют полисинаптические рефлексы. Элементарный рефлекс спинного мозга осуществляется по цепи из трех нейронов. Первый центростремительный чувствительный нейрон воспринимает раздражение: например, укол пальца своими окончаниями в коже и через тело клетки, находящееся в межпозвоночном ганглии, проводит по задним корешкам спинного мозга импульсы в задние рога его серого вещества, где находится второй промежуточный нейрон, передающий полученные сигналы в передние рога серого вещества. Здесь располагается тело третьего центробежного (двигательного) нейрона, который своим длинным концом, выходящим через передние корешки, вызывает сокращение мышц руки и она отдергивается.
Полисинаптические рефлексы могут возникать через посредство мышечных, кожных и висцеральных рецепторов. Они называются двигательными, если их эффекторами являются скелетные мышцы, вегетативными, если их рефлекторные дуги оканчиваются в на эффекторах вегетативной нервной системы.
Такого рода рефлексы играют существенную роль в движении (локомоторные рефлексы), питании (пищеварительные рефлексы), защите организма от опасных внешних воздействий (оборонительные рефлексы).
Если в рефлекторной дуге имеется два центральных синапса, то это дисинаптическая рефлекторная дуга. Дисинаптическая рефлекторная дуга обеспечивает сгибательный рефлекс. Если вызвать болевое раздражение задней конечности спинального животного (ущипнуть, ударить электротоком, прикоснуться горячим предметом), она отдергивается путем сгибания в голеностопном, коленном и бедренном суставах. Болевое раздражение передней конечности вызовет аналогичный сгибательный рефлекс. Соответсвующие рецепторы находятся в коже. Обеспечиваемое ими движение направлено на удаление конечности от источника болевого (а следовательно, повреждающего фактора); значит это типичный оборонительный рефлекс. Афферентные волокна образуют не только моносинаптические возбуждающие связи с гомонимными мотонейронами , но и тормозные связи с мотонейронами-антагонистами. Тормозная рефлекторная дуга включает центральный интернейрон, следовательно относится к дисинаптическому типу - в ней два центральных синапса: возбуждающий между афферентными волокнами и интернейронами и между аксонами интернейронов и мотонейронами.
Таким образом, афферентный нейрон через моносинаптическую связь оказывает возбуждение гомонимного мотонейрона , а через вставочный нейрон - тормозное влияние на мотонейроны антагонистов.
Еще один пример - перекрестный разгибательный рефлекс. Рефлекторное сгибание задней и передней конечности часто сопровождается разгибанием противоположной (контралатеральной) конечности, особенно при болевом раздражении. Импульсы с волокон, проводящих боль, переходят на противоположную сторону спинного мозга, запуская там реакцию разгибания.
В данном случае, болевое раздражение конечности на сегментарном уровне активирует четыре двигательные рефлекторные дуги: на ипсилатеральной стороне
- сгибатели возбуждаются,
- разгибатели тормозятся;
на контрлатеральной стороне
- сгибатели торомозятся,
- разгибатели возбуждаются.
Рефлекторные же дуги большинства таких рефлексов содержат более одного вставочного нейрона (часто очень много).
В дуге вегетативного рефлекса между афферентными и постганглионарными нейронами имеется по меньшей мере три синапса, два из которых лежат в сером веществе спинного мозга, а один - в вегетативном ганглии. В качестве примера можно привести кожно-висцеральный, висцеро-кожный, висцеро-соматический рефлексы.
Примером полисинаптического безусловного рефлекса может служить и сосательный рефлекс, который относится к пищевым. В его полисинаптической рефлекторной дуге рецепторами служат вопринимающие прикосновение структуры в коже губ (механорецепторы), а благодаря вставочным нейронам эффекторами являются мышцы губ, щек, языка, глотки, грудной клетки и диафрагмы. При этом сосательные движения должны координироваться с нормальными дыхательными движениями.
Если кусочек смоченной кислотой фильтровальной бумаги или ватки, положить на кожу спины декапитированной лягушки, то после короткого латентного периода животное сбросит бумажку (ватку) задней лапкой, которая окажется ближе к месту раздражения. Это пример оборонительного рефлекса. Его рецепторы находятся в коже спины, а мышцы конечности служат эффекторами.
Свойства полисинаптических рефлексов:
- при полисинаптических рефлексах подпороговые стимулы могут суммироваться до надпорогового. (Например, слабое “щекотание” или “царапание” в горле вызывает кашель не сразу, а через некоторое время. Такая суммация представляет собой центральное явление, т.е. оно происходит во вставочных нейронах и мотонейронах рефлекторной дуги.
- время рефлекса сильно зависит от интенсивности стимула - по мере увеличения интенсивности стимула (даже если она превысила порог) промежуток времени между началом стимула и завершением рефлекса (время рефлекса) становится короче - время рефлекса при рефлексе растяжения практически постоянно. Этот эффект обусловлен в основном временной и пространственной суммацией в центральных нейронах рефлекторной дуги благодаря усилению ритмического разряда рецепторов.
- форма ответа зависит от интенсивности стимула, т.е. при сильных стимулах рефлекс распространяется на эффекторы (например, группы мышц), которые ранее не участвовали в ответе.
- иррадиация. При сильных ститмулах возбуждение, которое вначале было подпороговым, переходит в надпороговое. Иррадиацию хорошо демонстрирует кашлевой рефлекс - слабое покашливание включает в основном мышцы самой глотки, тогда как при сильном кашле вступают мышцы грудной клетки, плеч, живота, диафрагмы.
- локальный знак - локализация стимула может влиять на ответ. Например, при болевом раздражении ноги сокращение флексорных мышц суставов бедра, колена и стопы зависят от места раздражения.
- привыкание (габитуация) - например, при ритмическом поглаживании живота в одном и том же месте с одной и той же интенсивностью рефлекс ослабевает, несмотря на то, что возбудимость участвующих рецепторов, мотонейронов и скелетных мышц не меняется. Изменение места или параметров раздражения (особенно увеличение его интенсивности) восстанавливает нормальный рефлекторный ответ. Такое восстановление называется отвыканием или дисгабитуацией. Возвращение к исходному ответу наблюдается также после перерыва раздражения на некоторое время.
Причину габитуации видят в синаптической депрессии.
- сенситизация - порог рефлекса снижается, время рефлекса укорачивается, рецептивное поле раширяется, и рефлекс иррадиирует. Пример - ритмические болевые стимулы.
- кондиционирование - длительное изменение рефлекторного ответа благодаря способности полисинаптических рефлексов адаптироваться в процессе научения.
Для спинномозговых рефлексов в общем характерна их относительная бедность, незначительность количества комбинаций деятельности разных эффекторов. Они нередко ограничены деятельностью эффекторов, иннервируемых от тех же сегментов мозга, где расположены рецепторы, раздражение которых вызывает данный рефлекс. Сокращение мышцы в ответ на ее растяжение, сгибание ноги кожа которой подвергается щипку или уколу, опорожнение мочевого пузыря в ответ на его растяжение, реакция мускулатуры прямой кишки при стимуляции ее рецепторов - вот типичные спинномозговые рефлексы. Правда, из преведенных выше примеров следует, что и здесь наблюдаются более сложные акты, такие, как перекрестный разгибательный рефлекс. Однако рефлекторная деятельность спинного мозга лишь в незначительной степени объединяет функции различных эффекторов.
При включении в рефлекторную деятельность продолговатого мозга отмечается существенное усложнение рефлекторной деятельности, что ведет к осуществлению таких рефлексов, как рвота, элементарная рефлекторная саморегуляция кровяного даления и дыхания в ответ на импульсы с рецепторов аортально-каротидной зоны, одновременное тоническое сокращение всех мышц-разгибателей, растягиваемых силой тяжести и др. В этих рефлексах проявляюся не только особенности рефлекторных дуг продолговатого мозга, связанного с высшими рецепторами, но и влияния, оказываемые продолговатым мозгом на весь спинной мозг.
При присоединении к рефлекторной деятельности спинного и продолговатого мозга деятельности красных ядер среднего мозга и филогенетически старых отделов мозжечка появляются новые рефлекторные акты - рефлексы выпрямления (у хищных и грызунов, но не у обезьян), торможение тонуса разгибателей, непомерно сильно у децеребрированных животных.
Включение в рефлекторную деятельность среднего мозга создает основу для осуществления таких рефлекторных актов, как восстановление определенной позы тела (“стояние”) при ее нарушении, отмечается некоторое изменение дыхательной деятельности.
С включением в рефлекторную деятельность промежуточного мозга (зрительных бугров и подбугровой области) происходит усложнение рефлекторной деятельности внутренних органов и ее объединение с двигательными рефлексами. Становится возможным осуществление таких рефлекторных актов, как теплорегуляции и реакции, обозначаемые термином “мнимое бешество”.
При включении в нервную деятельность все более высоких уровней центральной нервной системы рефлексы прогрессивно усложняются и видоизменяются.
Сложнейшие безусловные рефлексы (инстинкты). (лат. instictus - побуждение) - врожденная видоспецифическая форма поведения, становление которой завершается в постнатальном онтогенезе под влиянием и при взаимодействии с условными рефлексами. Безусловные рефлексы сложнейшие (инстинкты) - целостный поведенческий комплекс, системное морфофизиологическое образование, включающее в себя побуждающие и подкрепляющие компоненты.
Они характеризуются стереотипной видоспецифической последователь-ностью реализации поведенческого акта. Они возникают при первой их необхоимости, при появлении “специфического” для каждого из них раздражителя, обеспечивая тем самым неуклонность выполнения наиболее жизненно важных функций организма независимо от случайных, преходящих условий среды. Инстинктивные реакции носят врожденный характер и обладают высокой видовой специфичностью.
Особенности организации безусловного рефлекса сложнейшего. В реализации инстинктивных реакций пусковую функцию выполняют внешние стимулы. Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название “ключевые раздражители” или релизеры (разрешители). Каждый ключевой стимул запускает соответствующий ему комплекс стереотипных действий. Ключевые раздражители являются такими признаками внешней среды, на которые животные могут реагировать независимо от индивидуального опыта врожденным поведенческим актом. Пусковые стимулы как бы принудительно заставляют животное выполнять определенные комплексы действий, невзирая на воспринимаемую животным общую ситуацию.
Важную роль играют командные нейроны - клетки, активация которых запускает соответствующий поведенческий акт, однако сами не являются мотонейронами.
В последние годы сложилась общая схема организации инстинктивного поведения (рис. ). По этой схеме ключевой раздражитель может запускать соответствующую ему программу поведенченского акта на основе “жестких”, генетически детерминированных синаптических связей между сенсорными (чувствительными) и двигательными системами. Такой поведенческий акт осуществляется вне зависимости от общей внешней ситуации.


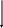

 Система
Система
[



 C]
р
Сенсорная
запуска,
Моторная
[П]
C]
р
Сенсорная
запуска,
Моторная
[П]
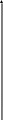 система
контроля система
система
контроля система

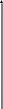 поведения
поведения


Модулирующая
система
С - стимул, Р- рецепция, П - поведенческий акт, пунктирная линия - модулирующее влияние, сплошная - деятельность модулирующей системы как оценочной инстанции.
Вместе с тем в организации сложного инстинктивного поведения значительную роль может играть перераспределение приоритетов рекций. Актуализированная (доминирующая) потребность и возникщее на ее основе мотивационное возбуждение повышают чувствительность сенсорных систем, избирательно настроенных на внешние стимулы, адекватные данной потребности. При этом осуществляется селективная (избирательная) активация нервных центров, связанных с формированием и запуском определенных программ двигательной активности, направленных на поиск ключевого раздражителя.
В результате селективной настройки афферентных, центральных и эфферентных звеньев центральной нервной системы появление стимула, адекватного доминирующей потребности, становится эффективным для запуска стереотипного рефлекторного акта.
Однако в структуре сложного инстинктивного поведения, кроме стереотипных реакций, выделяют еще и пластичные формы реагирования. Иными словами, в структуре инстинктов предусмотрены механизмы внутренней активности, “собственной инициативы” выбора поведения, которая проявляется прежде всего в поиске животным необходимых пусковых ситуаций и выборе наиболее эффективных способов для выполнения поведенческого акта. В инстинктивном поведении выделяют поисковую (пластичную) и завершающую (ригидную) фазы (по У.Крэгу) или подготовительную и исполнительную деятельность (по Ю. Конорскому).
Рассмотрим в качестве примера пищевой инстинкт. Дефицит питательных веществ в крови активирует вначале систему дорсального гиппокампа, ретикулярную формацию ствола мозга и неспецифические ядра таламуса (на рис. этот путь обозначен точками). Происходит неспецифическая активация подкорковых и корковых структур мозга. Затем приводятся в действие специфические механизмы положительной пищевой мотивации, главным образом медиального гипоталамуса, миндалин и коры, обусловливающие пищевое поведение (путь обозначен штриховкой). Поедание пищи и насыщение связано с деятельностью соответствующих стволовых, таламических и корковых структур (пути, обозначенные прерывистыми линиями). Когда насыщение достигнуто, в медиальных ядрах гипоталамуса активируются элементы отрицательной пищевой мотивации (зачернены) и еда прекращается. Участие коры мозга в этих последовательных этапах пищевого поведения представлены в виде смены соответствующих обозначений (точки, штриховка, прерывистая линия, зачернение).
Инстинкты очень часто осуществляются как цепные рефлексы, в которых осуществление одного звена рефлекторного акта ведет к раздражению рецепторов, вызывающему следующее звено этого акта. Так, например, у ряда животных определенные сезонные изменения в природе. воздействуя на ряд рецепторов, вызывает половой инстинкт, который после осуществления ряда включенных в него звеньев (например, различных голосовых и двигательных реакций) заканчивается процессами, ведущими к оплодотворению. Далее, раздражения интерорецепторов, связанные с беременностью или яйцеклеткой, часто в свою очередь ведут к комплексу различных рефлекторных актов, например, к высижыванию яиц. Точно так же рождение детеныщей, вылупливание птенцов вызывает новые рефлекторные акты родителей, ведущие к питанию потомства и пр. Важным звеном в цепи рефлекторных реакций, входящих в рефлекторный акт, часто является изменение выработки гормонов железами внутренней секреции.
Выделяют несколько групп рефлексов данного типа:
1) Витальные - обеспечивают индивидуальное и видовое сохранение организма. Сюда относятся пищевой, питьевой, регуляции сна, оборонительный и ориентировочный рефлексы, рефлекс экономии сил и многие другие. Критерием рефлексов витальной группы являются следующие:
- неудовлетворение соответствующей потребности ведет к физической
гибели особи,
- реализация рефлекса не требует участия другой особи того же вида.
2) Ролевые (зоосоциальные) - могут быть реализованы только путем взаимодействия с другими особями своего вида. Эти рефлексы лежат в основе полового, родительского, территориального поведения, в основе феномена эмоционального резонанса (“сопереживания”) и формирования групповой иерархии, где отдельная особь неизменно выступает в роли брачного партнера, родителя или детеныша, хозяина территории или пришельца, лидера или ведомого.
3) Безусловные сложнейшие рефлексы саморазвития - ориентированы на освоение новых сред, обращены к будущему. Они не связаны с индивидуальной и видовой адаптацией к наличной, в данный момент существующей ситуации. В эту группу рефлексов включают исследовательское поведение, безусловный рефлекс сопротивления (свободы), имитационный (подражательный) и игровой.
Особенностью группы безусловных рефлексов саморазвития является их самостоятельность; она не выводима из других потребностей организма и не сводится к другим мотивациям. Так, реакция преодоления преграды осуществляется независимо от того, какая потребность первично инициировала поведение и какова цель, на которой возникла преграда. Это в данном случае не имеет значения. Именно характер преграды (стимульно-преградная ситуация), а не первичный мотив определяет состав действий в поведении, которое способно привести к цели.
Подводя итог, следует отметить, что сложнейшие безусловные рефлексы представляют собой комплекс включающихся в определенной последовательности безусловных рефлексов (т.е. цепной характер реакции, когда завершение одного рефлекса, служит сигналом следующего рефлекса), при жестком объединении участвующих в их осуществлении нервных структур, а непосредственным сигналом к его включению служит конкретный раздражитель. Важной характеристикой является и то, что они зависят от гормональных и метаболических факторов, кроме того они характеризуются многими свойствами доминанты.
УСЛОВНЫЕ РЕФЛЕКСЫ. Вперые выделив такое явление как условный рефлекс, И.П.Павлов увидел в нем высшую форму рефлекторной деятельности - реакцию не на сам раздражитель, а на сигнал, предшествующий этому раздражителю. Реакция человека и животного на сигнал имеет то преимущество, что позволяет избежать действие раздражителя, если он отрицателен (опасен), или поспешить навстречу этому раздражителю, если он положителен (необходим или приятен).
В естественных условиях безусловные рефлексы, с которыми рождается живое существо, в результате взаимодействия с многочисленными изменчивыми факторами внешней среды как бы “обрастают” разнообразными условными рефлексами и фактически перестают существовать в чистом виде. Так, например, безусловный рефлекс - выделение слюны при попадании пищи в полость рта, преобразуется: слюна выделяется на вид пищи, ее запах, на упоминание о пище в разговоре (см. М.Е.Салтыков-Щедрин “Как один мужик двух генералов прокормил”) и т.д. Такие условные рефлексы, формирующие качество самого безусловного рефлекса, воспринимаемые не вкусовым анализатором, а другими анализаторами (зрительным, обонятельным), называются натуральными или естественными, например, слюноотделение на вид пищи. Искусственными называются условные рефлексы, вырабатываемые на посторонние по отношению к безусловному раздражителю сигналы, например, слюноотделение на звук метронома.
Условные рефлексы бывают различные. Единой классификации не существует. В основе различных классификаций лежат разные признаки:
1. Локализация отдельных элементов рефлекторной дуги:
Рецепторное поле
- интерорецептивные-
-экстерорецептивные- УСЛОВНЫЕ РЕФЛЕКСЫ
-проприорецептивные
Центральное звено
- спинальные
- бульбарные
- мезэнцефалические
- мозжечковые
- диэнцефалические
- корковые
Эфферентное звено
- соматические
-вегетативные
2. Эффекторные изменения (по эффектору)
- глотательный
- мигательный
-кашлевой
и др.
3. Характер влияния на деятельность эффектора
- возбудимые
- тормозные
4. Генетическая предрасположенность
- безусловные
- условные
5. Биологическое значение ответной реакции
- пищевые
- оборонительные
- половые
- родительские условные рефлексы
6. Совпадение во времени сигнала и подкрепления
- совпадающие - сразу после включения сигнала дают подкрепление
- отставленные - подкрепление подается к сигналу лишь через
некоторое время (обычно в пределах 5-30 с)
- запаздывающие - время отставания удлиняется до 1-3 мин
- следовые - подкрепление вводится после выключения сигнала
7. Сложность (или уровень интеграции)
- первого порядка
- второго порядка
- третьего порядка
и т.д.
Рефлекс первого порядка - предполагает наличие одного раздражителя и одного сигнала: например свет-пища, в результате на свет начинает выделяться слюна.
Рефлексы первого порядка могут служить базой для выработки рефлексов второго порядка - два последовательных сигнала и один раздражитель. Применяется дополнительный сигнал, например звук, предшествующий световому сигналу. В результате многократного сочетания звук начинает вызывать слюноотделение. Т.е. звук приводит к возбуждению центра зрения, а последний возбуждает пищевой центр.
Рефлексы третьего порядка. При достаточно прочном условном рефлексе второго порядка можно приступить к выработке рефлекса третьего порядка. Для этого используется новый раздражитель (сигнал), например, прикосновение к коже с помощью специального прибора, называемого касалкой. В этом случае касание подкрепляется звуком, звук приводит к возбуждению центра зрения, а последний возбуждает пищевой центр. Таким образом вырабатывается еще более сложная опосредованная связь.
При повышенном мотивированном возбуждении у собак удается выработать условные рефлексы 2-4 порядков.
Рефлексы более сложного, высокого порядка (4,5,6 и т.д) вырабатываются только у приматов и человека. Существуют данные о том, что у взрослых людей вырабатываются условные рефлексы от 2-го до 20-го порядков. Однако, условные рефлексы высоких порядков нестойкие и быстро угасают.
Таким образом, сложность рефлекса определяется количеством использованных сигналов.