

Вопрос №139
Гормоны, регулирующие обмен кальция и фосфора: паратгормон, кальцитонин, калитриол. Структура, регуляция. Нарушения - гипо- и гиперпаратиреоз.
Нормальная концентрация Са2+ в крови - 2,12-2,62 мМ/л.
В организме содержится 100-1200 г Са2+:
99% - в минерализованных тканях (костях, зубах) в виде кристаллов гидроксиапатита.
Остальная часть (1%) - в крови:
50% (0,5% от всего Са2+) - в свободном виде
50% (0,5% от всего Са2+) - связаны с альбумином и цитратом.
Функции Са2+ :
основной компонент минерализованных тканей
второй посредник действия гормонов
кофактор ферментов (α-амилаза)
участвует в мышечном сокращении
участвует в свертывании крови (IV фактор).
Гормоны:
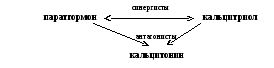
Паратгормон
Состоит из 84 аминокислотных остатков.
Синтезируется в клетках паращитовидной железы.
Действует через цАМФ.
Повышает концентрацию Са2+ в крови.
Органы мишени:
костная ткань - вызывает резорбцию костной ткани.
Клетки-мишени - остеобласты. Они синтезируют биологически активные вещества - цитокины, которые воздействуют на остеобласты и активируют их, а остеокласты резорбируют кость. Следовательно, Са2+ выходит в кровь;
почки
усиливает реабсорбцию Са2+ и выведение фосфор
активизирует синтез своего синергиста - кальцитриола
Кальцитриол
Производное холестерола.
Рецепторы в цитоплазме.
Вторых посредников нет.
В коже под УФ холестерол превращается в витамин D3 - холекальциферол. Он поступает в печень, где гидроксилируется (присоединение ОН-группы в положении 25). Затем поступает в почки, где гидроксилируется по 1 положению. Образуется 1,25-дигидроксихолекальциферол = кальцитриол.
Органы-мишени:
кишечник (в основном) - гормон стимулирует синтез белков-переносчиков для Са2+ . Следовательно, улучшается всасывание Са2+ ;
кости - активирует остеокласты.
Кальцитонин
Состоит из 32 аминокислотных остатков.
Вырабатывается в парафолликулярных клетках щитовидной железы.
Действует через цАМФ.
Понижает концентрацию Са2+ в крови.
Органы-мишени:
кости - активирует остеобласты, включает Са2+ в кости, способствуя минерализации;
почки - усиливает выведение Са2+ с мочой.
Вопрос №140
Производные арахидоновой кислоты - эйкозаноиды. Представители, их синтез и биологическая роль.
Эйкозаноиды - это большая группа веществ, которые могут синтезироваться почти всеми типами клеток, за исключением эритроцитов, и как гормоны местного действия оказывают эффекты по паракринному или аутокринному механизму через специфические рецепторы.
Главные биологические эффекты эйкозаноидов:
• участвуют в регуляции сокращений гладкой мускулатуры (разные типы эйкозаноидов вызывают вазоконстрикцию или вазодилатацию, бронхоконстрикцию или бронходилатацию);
• регулируют экскрецию воды и Na+ почками и артериальное давление;
• участвуют в развитии воспаления;
• регулируют свертываемость крови
Основные классы эйкозаноидов представлены:
• простагландинами PG (включая простациклины);
• тромбоксанами;
• лейкотриенами.
Исходными субстратами для синтеза эйкозаноидов являются полиеновые жирные кислоты с 20 атомами углерода. Главный субстрат для синтеза эйкозаноидов у человека - арахидоновая кислота (20:4 ω-6).
Полиеновые кислоты с 20 атомами углерода поступают в организм человека с пищей или образуются из незаменимых жирных кислот с 18 атомами углерода, также поступающих с пищей.
В разных тканях из арахидоновой кислоты под действием специфического для этой ткани набора ферментов образуются различные эйкозаноиды. Обычно в каждом типе клеток синтезируется преимущественно один тип эйкозаноидов.
Образовавшиеся в клетке эйкозаноиды выходят из нее и взаимодействуют с рецепторами на поверхности этой же клетки (аутокринный механизм) или с рецепторами на соседних клетках (паракринный механизм). Время полураспада простагландинов равно нескольким минутам. Для каждого эйкозаноида есть несколько типов рецепторов, которые располагаются в мембране клеток рядом с аденилатциклазой, некоторые из простагландинов взаимодействуют с G-белками аденилатциклазной системы. Такие простагландины, взаимодействуя со своими рецепторами, могут модулировать активность аденилатциклазы. Ответ клетки на действие эйкозаноидов определяется типом ее рецепторов.
С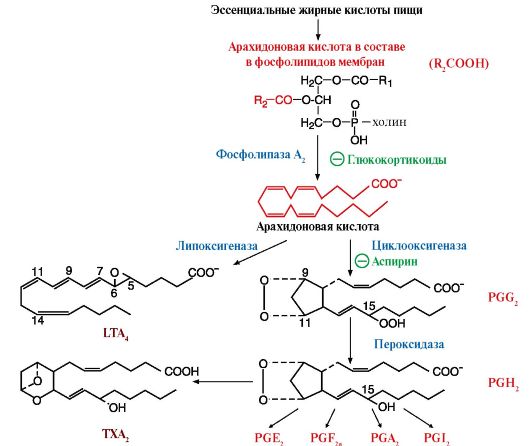 интез
эйкозаноидов начинается после отделения
жирной кислоты от фосфолипидов мембран
под действием фермента фосфолипазы A2.
Арахидоновая кислота переходит в
цитозоль клетки и становится доступной
для синтеза эйкозаноидов. Синтез основной
группы эйкозаноидов - простагландинов,
простациклинов и тромбоксанов - начинается
с действия на полиеновую кислоту фермента
- простагландинсинтазы.
интез
эйкозаноидов начинается после отделения
жирной кислоты от фосфолипидов мембран
под действием фермента фосфолипазы A2.
Арахидоновая кислота переходит в
цитозоль клетки и становится доступной
для синтеза эйкозаноидов. Синтез основной
группы эйкозаноидов - простагландинов,
простациклинов и тромбоксанов - начинается
с действия на полиеновую кислоту фермента
- простагландинсинтазы.
Если арахидоновая кислота подвергается действию другого фермента - липоксигеназы, то образуются молекулы с тремя сопряженными двойными связями (отсюда название «лейкотриены»). Они имеют несколько вариантов структур и в основном участвуют в развитии аллергических реакций. Синтез большинства эйкозаноидов увеличивается при воспалительных процессах. Активность фосфолипазы А2 при этих состояниях повышается и субстраты становятся доступными для синтеза эйкозаноидов.
Ингибиторами синтеза эйкозаноидов являются:
- глюкокортикоиды, которые индуцируют синтез группы белков, ингибирующих активность фосфолипазы А2, и таким образом подавляют синтез всех типов эйкозаноидов, участвующих в воспалении. Эти препараты обладают сильным противовоспалительным свойством.
- аспирин и другие нестероидные противовоспалительные препараты необратимо ингибируют циклооксигеназу.
Роль эйкозаноидов в регуляции свертывания крови. В норме свертывающая и противосвертывающая системы крови пребывают в состоянии равновесия, при котором кровь находится в жидком состоянии, но способна быстро образовывать тромб при возникновении соответствующих условий. При патологии или при действии фармакологических средств это равновесие может смещаться в любую сторону. В норме клетки эндотелия сосудов продуцируют простациклины PGI2, PGE2, PGD2, которые препятствуют агрегации тромбоцитов и сужению сосуда; TXA2, стимулирующий агрегацию тромбоцитов, в этих условиях не секретируется. ТХА2 секретируется тромбоцитами только в результате их активации - например, при контакте с поврежденной стенкой кровеносного сосуда. При разрушении клеток эндотелия (например, в результате образования атеросклеротической бляшки) синтез PGI, PGE, PGD снижается. Тромбоциты активируются в месте контакта с поврежденной стенкой сосуда и секретируют ТХА2, что стимулирует образование тромба в области повреждения эндотелия сосудов и развитие инфаркта.
Если с пищей поступает больше жирных кислот ω-3, которые в большом количестве содержатся в рыбьем жире, то эти кислоты включаются преимущественно в фосфолипиды мембран (вместо арахидоновой) и после действия фосфолипазы А2 являются основными субстратами для синтеза
В норме клетки эндотелия продуцируют PGI2, который вызывает релаксацию гладкой мускулатуры сосудов и ингибирует агрегацию тромбоцитов. Тромбоциты в неактивном состоянии не продуцируют тромбоксаны, кровь находится в жидком состоянии. NO - оксид азота, продуцируемый ферментом NO-синтазой, также обладает вазодилятаторным эффектом эйкозаноидов.
При поражении клеток эндотелия сосуда (например, в результате развития атеросклеротической бляшки) синтез PGI2 в данном участке стенки сосуда не происходит. В это время тромбоциты контактируют с поврежденной сосудистой стенкой, в них активируется фосфолипаза А2, освобождается арахидоновая кислота и из нее синтезируется ТХА2. ТХА2 стимулирует агрегацию тромбоцитов и сокращение стенок сосуда, в результате чего на поврежденном участке сосуда образуется тромб, происходит резкое сужение просвета сосуда, нарушается кровоснабжение ткани и может развиться инфаркт